
Abstrak
Keluarga faktor transkripsi E-Twenty-Six (ETS) yang dilestarikan secara evolusioner bertindak di hilir jalur transduksi sinyal utama dan memainkan peran penting dalam pengembangan dan pemeliharaan jaringan. Yang penting, fungsinya sering kali rusak dalam sebagian besar jenis tumor, dan mereka juga sangat diperlukan untuk perkecambahan angiogenik, ciri khas kanker, yang penting untuk memicu pembesaran dan penyebaran tumor. Akibatnya, menargetkan aktivitas ETS yang tidak normal berpotensi mewakili cara yang tepat dan efektif untuk memblokir pertumbuhan tumor. Di sini, kami menyajikan penyaringan throughput tinggi bukti prinsip dan karakterisasi awal dari kandidat hit, sebagai kerangka metodologis dan konseptual untuk mengidentifikasi penghambat faktor transkripsi ETS baru, yang pada akhirnya dapat mengarah pada jalan terapeutik baru untuk mengobati kanker.
Singkatan
AP1 , protein pengaktif 1
DLAV , pembuluh anastomosis longitudinal dorsal
DTT , ditiotreitol
EDBD , domain pengikat DNA ETS
ETS , E-dua puluh enam
GFP , protein fluoresensi hijau
hiPSC-CM , kardiomiosit turunan sel induk pluripoten yang diinduksi manusia
HTRF , fluoresensi waktu-terselesaikan homogen
HTS , layar throughput tinggi
LCMS , kromatografi cair spektrometri massa
NMR , resonansi magnetik nuklir
PBS , garam penyangga fosfat
PECAM1 , molekul adhesi trombosit dan sel endotel 1
PH , homologi plekstrin
RFU , unit fluoresensi relatif
STR , pengulangan tandem pendek
TERT , telomerase transkriptase balik
TINS , target penyaringan NMR yang diimobilisasi
VEGF , faktor pertumbuhan endotel vaskular
Dalam dua dekade terakhir, perubahan besar dalam pendekatan farmakologis untuk pengobatan kanker telah melibatkan peralihan dari kemoterapi tradisional menuju terapi presisi yang diarahkan terhadap protein spesifik dalam sel tumor (dan stroma tumor) dengan tujuan membatasi sitotoksisitas pada jaringan sehat [ [ 1 – 5 ] ]. Meskipun ada peningkatan dalam hasil pasien, revolusi yang dijanjikan sejauh ini gagal terwujud. Dua alasan utama untuk ini adalah perolehan resistensi terhadap pengobatan [ [ 6 – 10 ] ] dan fakta bahwa kategori inhibitor ini, hingga saat ini, telah menargetkan rentang protein yang relatif terbatas yang terdiri dari kinase dan reseptor permukaan sel [ [ 11 – 13 ] ]. Salah satu solusi potensial untuk masalah ini adalah mengidentifikasi target baru, terutama sekarang karena ada penerimaan yang lebih luas tentang kegunaan pencarian di luar batasan historis dari genom yang dapat diberi obat sebelumnya (pada dasarnya terdiri dari enzim yang disebutkan di atas, dan protein permukaan sel) [ [ 14 , 15 ] ].
Dalam konteks ini, Darnell [ [ 16 ] ] benar untuk menyatakan bahwa ada sejumlah terbatas faktor transkripsi yang terlalu aktif dalam kanker, yang dapat menjadi kandidat yang cocok untuk pengembangan obat antikanker. Dengan demikian, ada alasan kuat untuk berpikir bahwa keluarga faktor transkripsi ETS dapat mewakili target yang sangat baik karena anggota keluarga ETS dapat memainkan peran penting dalam mengatur banyak proses penting dalam biologi sel dan jaringan kanker termasuk kontrol siklus sel, proliferasi sel, diferensiasi sel dan metastasis [ [ 17 – 19 ] ]. Anggota pendiri, dan prototipe, dari keluarga, ETS1, pertama kali diidentifikasi sebagai protein fusi onkogenik dalam retrovirus leukemia burung E26 [ [ 20 , 21 ] ]. Sampai saat ini, 28 anggota keluarga ETS yang berbeda telah dideskripsikan pada manusia, yang diklasifikasikan menjadi 12 subkelompok berdasarkan homologi urutan domain protein [ [ 22 ] ]. Secara operasional, protein ETS berfungsi dengan mengikat situs DNA konsensus (meliputi urutan inti GGA(A/T)) melalui domain pengikatan DNA ETS (EDBD) yang sangat terkonservasi, struktur heliks-putar-heliks bersayap (sekitar) 85 asam amino yang dikodekan oleh semua anggota keluarga [ [ 22 , 23 ] ]. Dalam 40 tahun terakhir, banyak bukti telah mengungkap pentingnya faktor ETS dalam evolusi banyak tumor hematologi dan padat [ [ 24 , 25 ] ]. Beberapa mekanisme berbeda telah terbukti mendasari misregulasi ETS patologis termasuk penataan ulang kromosom, yang menghasilkan fusi gen ETS pada leukemia, sarkoma Ewing, kanker payudara, kanker lambung, kanker kepala dan leher, dan kanker prostat; amplifikasi gen pada kanker payudara dan melanoma; mekanisme perolehan fungsi yang terkait dengan peningkatan aktivitas dan stabilitas faktor ETS, misalnya, pada glioblastoma, melanoma, dan kanker kandung kemih [ [ 26 ] ]. Selain itu, survei genomik skala besar mengidentifikasi mutasi pada gen telomerase reverse transcriptase (TERT), yang mengkode subunit telomerase reverse transcriptase katalitik, sebagai salah satu perubahan genetik yang paling umum dalam berbagai macam tumor [ [ 27 , 28 ] ]. Sementara aktivitas telomerase biasanya tidak ada dalam sel somatik postembrionik, ia diaktifkan kembali dalam 90% kanker agresif melalui peningkatan regulasi TERT [ [ 29 , 30 ] ]. Dua mutasi yang saling eksklusif dekat dengan situs awal transkripsi, yang menciptakan de novoSitus pengikatan ETS dan diduga mengarah pada pengikatan abnormal faktor ETS tertentu, diyakini mendasari peningkatan regulasi aktivitas TERT yang mendorong tumor [ [ 27 , 28 ] ]. Yang penting, faktor transkripsi ETS juga sangat diperlukan untuk perkecambahan angiogenik [ [ 31 – 33 ] ], termasuk proses di mana tumor merangsang perluasan pembuluh darah lokal, yang secara fisik menghubungkannya ke sirkulasi, sehingga memicu pertumbuhan dan penyebaran tumor.
Mengingat alasan yang kuat untuk menargetkan protein ETS sebagai terapi antikanker yang potensial [ [ 34 , 35 ] ], dalam beberapa tahun terakhir, metode kimia komputasional dan penyaringan fenotipik dan fungsional telah digunakan untuk menemukan inhibitor ETS [ [ 36 – 42 ] ]. Namun, ada tantangan teknis yang harus diatasi untuk mengidentifikasi inhibitor spesifik yang secara langsung memblokir aktivitas ETS, paling tidak karena faktor transkripsi diperkaya dengan daerah yang tidak teratur secara intrinsik dan tidak memiliki kantong pengikat yang jelas dan terdefinisi dengan baik [ [ 43 , 44 ] ], yang menjelaskan sedikitnya kampanye penyaringan molekul kecil yang berhasil hingga saat ini. Meskipun ada kendala yang mungkin seperti itu, EDBD berpotensi cukup terstruktur untuk berinteraksi dengan molekul kecil, dan karena aktivitas ETS sebagian besar bergantung pada pengikatan DNA melalui EDBD, domain ini dapat mewakili target yang dapat ditangani. Dalam penelitian ini, kami menggunakan metodologi yang dioptimalkan untuk menargetkan EDBD. Dua strategi utama telah digunakan. Pertama, kami menggunakan target immobilized nuclear magnetic resonance (NMR) screening (TINS) [ [ 45 ] ] untuk mengidentifikasi perancah molekul kecil yang dapat mengikat langsung ke EDBD. Mengingat skala yang relatif lebih kecil, pendekatan bukti prinsip tersebut telah digunakan secara rutin oleh industri farmasi/bioteknologi sebagai langkah awal untuk menguji potensi ligandabilitas protein target [ [ 46 , 47 ] ]. Pendekatan kedua menggunakan penyaringan fungsional multikomponen baru dengan throughput tinggi dari molekul kecil, yang mengganggu pengikatan ETS ke situs pengikatan DNA konsensus.
Pekerjaan kami menunjukkan bahwa pendekatan penyaringan berthroughput tinggi dapat digunakan untuk mengidentifikasi penghambat molekul kecil spesifik dari faktor transkripsi ETS.
Bahan dan metode
Ekspresi dan pemurnian protein
C-terminal His 6 epitope-tagged EDBD dikloning ke dalam pET28a dan diekspresikan dalam Rosetta DE3 Escherichia coli . Sel yang dipanen disuspensikan kembali dalam buffer lisis [500 mm NaCl , 20 mm imidazole , 2 mm DTT , 10% gliserol, 1× phosphate buffered saline (PBS), pH 7,4] di atas es dan disonikasi (30 detik aktif, 30 detik istirahat) selama 5 menit. Lisat disentrifugasi pada 24.500 x g selama 40 menit pada suhu 4 °C. Supernatan disaring melalui filter pengikat protein rendah 0,22-μm (Merck Millipore, Tullagreen, Cork, Irlandia), dimuat ke kolom HisTrap HP 5 mL (Cytiva, Uppsala, Swedia) dan dielusi dengan gradien dalam buffer elusi [500 m m NaCl, 300 m m imidazol, 2 m m dithiothreitol (DTT), 10% gliserol, 1× PBS, pH 7,4]. Fraksi yang dikumpulkan diencerkan 10 kali dalam buffer kolom heparin (20 m m HEPES, 1 m m DTT, pH 7,4), dimuat ke kolom HiTrap Heparin 5 mL (Cytiva) dan dielusi dengan gradien dalam Buffer Heparin B: 1 m NaCl, 20 m m HEPES, 1 m m DTT, pH 7,4. Fraksi protein dipekatkan menggunakan Amicon-Ultra Centrifugal Filter (Merck Millipore), dimasukkan ke dalam kolom filtrasi gel Superdex 200 16/600 (Cytiva) dan diseimbangkan dalam buffer (150 m m NaCl, 1 m m DTT, 1× PBS, pH 7,4). Fraksi dari setiap langkah dijalankan pada gel SDS/PAGE 12% dan diwarnai dengan Colloidal Blue Staining Kit Invitrogen (Thermo Fisher Scientific, Carlsbad, CA, AS).
Pemindaian NMR yang diimobilisasi target (TINS)
His C-terminal murni yang ditandai dengan 6 epitop ETV6 DBD [asam amino 1–118 (penomoran PDB 2DAO ) yang sesuai dengan asam amino 334–452] diimobilisasi melalui penggandengan amina ke 600 μL resin Actigel-ALD (Sterogene, Carlsbad, CA, AS) dalam 25 m m HEPES pH 7,5, 100 m m NaCl, dan 2 m m MgCl 2 pada suhu 4 °C (mengikuti protokol pabrik). Protein referensi [domain homologi pleckstrin (PH) dari Akt, yang merupakan permukaan protein khas tetapi tidak memiliki situs pengikatan molekul kecil tertentu] diimobilisasi menggunakan protokol yang sama. Efisiensi imobilisasi di atas 90%, dan konsentrasi akhir target yang diimobilisasi berada dalam kisaran 100 μ m . Percobaan TINS dilakukan pada spektrometer Bruker NMR 500 MHz menggunakan urutan pulsa Hadamard selektif spasial dan dianalisis seperti yang dijelaskan sebelumnya [ [ 46 – 49 ] ]. Pustaka fragmen terdiri dari 1364 senyawa dengan berat molekul rata-rata ~ 200 Da dan mematuhi aturan umum 3 (berat molekul fragmen < 300, cLogP ≤ 3, jumlah donor ikatan hidrogen ≤ 3 dan jumlah akseptor ikatan hidrogen ≤ 3, jumlah ikatan yang dapat diputar ≤ 3). Pustaka menunjukkan keragaman struktural yang signifikan, meliputi berbagai macam gugus fungsi, konfigurasi stereokimia, dan perancah yang berbeda. Pustaka dioptimalkan untuk menyaring ruang kimia yang sangat beragam dan mencakup kerangka aromatik, heterosiklik, dan alifatik dengan gugus fungsi seperti sulfonamida, amida, alkohol, keton, dan halida. Campuran fragmen, yang mengandung 2–8 senyawa per campuran, disiapkan dengan mencampur stok d6DMSO (pada 100 m m ) dan pengenceran lebih lanjut ke dalam buffer NMR (PBS terdeuterasi) hingga konsentrasi akhir 500 μ m.Campuran telah dirancang dengan cermat berdasarkan spektrum NMR 1D 1H individual dari fragmen untuk meminimalkan tumpang tindih spektral dan untuk menghindari masalah kelarutan. Penyaringan dilakukan dengan siklus berulang injeksi campuran fragmen ke dalam kedua sel sel ganda yang dikemas dengan EDBD yang diimobilisasi dan protein referensi. Setelah injeksi campuran, aliran dihentikan dan data NMR diperoleh. Fragmen dicuci sebelum injeksi berikutnya. Pengikatan fragmen ke EDBD dapat dideteksi dengan pengurangan sederhana pada tinggi sinyal NMR dari fragmen itu di hadapan protein target relatif terhadap intensitas puncak di hadapan protein referensi. Pengikatan preferensial fragmen ke protein target (T) dan referensi (R) (didefinisikan sebagai nilai T/R) ditentukan dengan menggunakan amplitudo sinyal senyawa di hadapan target dibagi dengan amplitudo sinyal senyawa di hadapan referensi (nilai T/R). Nilai batas T/R sebesar 0,5 dipilih untuk pemilihan kandidat hit. Pemindaian fragmen mengidentifikasi 103 kandidat hit unik (tingkat hit 7,5%). Layar molekul kecil dengan throughput tinggi Uji fluoresensi waktu-terurai homogen (HTRF) digambarkan secara skematis pada Gambar 3 dan dijelaskan dalam teks. Skrining dilakukan dalam format 1536-well (Corning, Kennebunk, ME, AS) (volume akhir 4 μL) menggunakan BioRAPTR (Beckman Coulter, Brea, CA, AS). Kondisi berikut dioptimalkan secara ketat: protein 40 nm /antibodi Eu-anti-His 4 nm (Cisbio Bioassays, Codolet, Prancis); Strep-XL665 62,5 nm (Cisbio)/oligo terbiotinilasi 62,5 nm ±500 nm oligo tak terbiotinilasi. Senyawa diuji pada konsentrasi 10 μm ( skor Z ′ = 0,76; kisaran 0,57–0,85). Pesaing oligo yang tidak terbiotinilasi dianggap sebagai penghambat maksimal, dan batas penghambatan 50% digunakan dalam uji primer untuk memilih senyawa potensial yang terkena. Sekitar 400.000 senyawa disaring, yang mana 398.892 dapat divalidasi dan diverifikasi. Sebanyak 2.706 senyawa dipilih untuk validasi lebih lanjut. Sebanyak 277 senyawa dapat dikonfirmasi dalam pengulangan penyaringan yang independen, yang mana 83 senyawa memberikan penghambatan >50% dan dipilih untuk karakterisasi lebih lanjut. Potensi dan efikasi dinilai menggunakan kurva dosis-respons dengan tujuh konsentrasi (kisaran 20 μm hingga 20 nm , n = 2), dengan kriteria pemilihan ditetapkan pada pEC50 > 5. Uji deseleksi menggunakan domain pengikatan DNA SMAD3 digunakan untuk menghilangkan senyawa yang menunjukkan aktivitas nonspesifik. Terakhir, analisis keminformatika menggunakan model Bayesian membantu menyempurnakan daftar dan selanjutnya menghilangkan kemungkinan positif palsu.
Penetapan urutan tulang punggung NMR
Escherichia coli BL21 (DE3) yang mengekspresikan His 6 epitope-tagged ETV6 EDBD dikultur dalam medium minimal M9 yang dilengkapi dengan 15 NH 4 Cl dan 13 C- d -Glukosa (CIL) sebagai satu-satunya sumber nitrogen dan karbon. Protein berlabel dimurnikan seperti dijelaskan di atas. Penugasan tulang punggung berurutan diperoleh dengan korelasi pergeseran kimia Cα dan Cβ dari residu i dan i-1 ke resonansi amida 1 H dan 15 N menggunakan spektrum HNCACB, CBCAcoNH, HNCA dan HNcoCA yang diperoleh pada 296 K pada spektrometer NMR BRUKER DMX 600 MHz yang dilengkapi dengan krioprobe TXI. Data yang diperoleh diproses menggunakan nmrpipe dan divisualisasikan pada Sparky [ [ 50 , 51 ] ]. Proses penugasan dipandu oleh pergeseran kimia yang diprediksi yang dihitung oleh SHIFTX dalam program penugasan otomatis MARS menggunakan struktur PDB- 2DAO sebagai referensi [ [ 52 , 53 ] ].
Analisis gangguan pergeseran kimia
Spektrum [ 1 H, 15 N]-HSQC diperoleh pada 296 K pada spektrometer NMR Bruker DMX 600 MHz yang dilengkapi dengan krioprobe TXI. Sampel NMR disiapkan dalam 25 mm HEPES pH 7,5, 100 mm NaCl , 1 mm EDTA , dan 1 mm β -merkaptoetanol. Sampel NMR yang umum mengandung 0,130 mm protein dan fragmen pada berbagai konsentrasi (lihat bagian Hasil ) dan 5% d6-DMSO. pH sampel disesuaikan dengan hati-hati dalam ±0,05 unit setelah penambahan senyawa. Sebanyak 128 peningkatan tidak langsung dengan 16 pemindaian per peningkatan diperoleh. Data diproses menggunakan Topspin 1.2/2.1 (Bruker, Billerica, MA, AS) dan divisualisasikan pada Sparky [ [ 51 ] ]. Gangguan pergeseran kimia dalam spektrum [ 1 H, 15 N]-HSQC dihitung berdasarkan perubahan nilai 1 H dan 15 N ppm dengan dan tanpa adanya senyawa (Δδ > dua kali simpangan baku + Δδavg). Situs pengikatan potensial suatu senyawa dipetakan ke permukaan struktur ETV6 EDBD (PDB- 2DAO ) menggunakan data gangguan pergeseran kimia dan dilihat dalam pymol (Schrödinger, New York, NY, AS).
Studi resonansi plasmon permukaan
Pengikatan DNA pada ETV6 EDBD diuji menggunakan resonansi plasmon permukaan pada instrumen biacore T200 (Cytiva). Pengintaian pH dilakukan untuk menentukan pH optimum untuk imobilisasi protein pada permukaan chip CM5. Setelah imobilisasi protein (6000 RU), peningkatan konsentrasi DNA dititrasi dalam mode kinetik siklus tunggal. Kondisi penyangga adalah sebagai berikut: 25 m m HEPES pH 7,5, 100 m m NaCl, 1 m m EDTA dan 1 m m β-merkaptoetanol. Analisis dilakukan menggunakan perangkat lunak evaluasi biacore (Cytiva) dan prisma graphpad (versi 10.2.3; Perangkat Lunak GraphPad, San Diego, CA, AS).
Kultur sel, biokimia
Lini sel kanker prostat, VCaP (RRID: CVCL_2235), dikulturkan dalam Iscove’s Modified Dulbecco’s Medium (IMDM) (Gibco, Grand Island, NY, AS), yang dilengkapi dengan 15% Fetal Bovine Serum (FBS) (Gibco). Sel sarkoma Ewing SK-ES-1 (RRID: CVCL_0627) dikulturkan dalam IMDM yang dilengkapi dengan 10% FBS. HUVEC primer (Lonza, Basel, Swiss) dikulturkan dalam medium EGM2 (Lonza). Sel MDA-MB-231 (RRID: CVCL_0062) dikulturkan dalam Dulbecco’s Modified Eagle Medium (DMEM) (Gibco) yang dilengkapi dengan 10% fetal bovine serum (FBS) (Gibco). Semua lini sel diperoleh dari American Type Culture Collection (ATCC, Manassas, VA, AS). Sel dipelihara dalam inkubator humidifikasi 5% CO2 , 37 °C, diuji setiap bulan untuk mengetahui adanya kontaminasi mikoplasma dan diperiksa keasliannya dalam 3 tahun terakhir oleh layanan internal menggunakan profil pengulangan tandem pendek (STR).
Pengujian proliferasi
Sel-sel disemai, dalam rangkap tiga, ke dalam pelat putih 96-sumur dengan dasar datar yang bening dalam 100 μL medium pada kepadatan awal berikut: VCaP, 30.000 sel·mL −1 ; HUVEC dan SK-ES-1, 40.000 sel·mL −1 . Medium tersebut dilengkapi dengan konsentrasi senyawa yang ditunjukkan (lihat bagian Hasil ). Jumlah sel yang hidup ditentukan menggunakan reagen Cell Titer-Blue (Promega, Madison, WI, AS) pada hari ke-0, ke-3, dan ke-5 setelah perlakuan. Pembacaan absorbansi dilakukan pada unit fluoresensi relatif (RFU) 544 nm/590 nm menggunakan VICTOR X3 Multilabel Plate Reader (PerkinElmer, Waltham, MA, AS).
Uji pengikatan DNA in vitro
Protein yang diterjemahkan secara in vitro dibuat menggunakan sistem transkripsi/translasi in vitro retikulosit yang digabungkan TNT (Promega). 50 pmol oligonukleotida untai ganda terbiotinilasi yang mengandung tiga situs pengikatan ETS konsensus (untuk faktor transkripsi ETV6, FLI1, ERG) atau tiga situs pengikatan protein aktivator-1 (AP-1) (untuk JUN) digabungkan ke manik-manik Dynabeads™ MyOne™ Streptavidin C1 (Invitrogen, Thermo Fisher Scientific). Oligonukleotida untai ganda diinkubasi dengan protein yang diterjemahkan secara in vitro dengan atau tanpa senyawa pada konsentrasi yang disorot dalam keterangan gambar. Reaksi diinkubasi pada suhu 4 °C, dengan pengocokan, selama 30 menit dengan adanya 1 μg poli (dI/dC), spermidin 4 m m . Manik-manik dicuci berturut-turut ×4 dengan buffer pengikat (50 mm KCl, 10 mm HEPES (pH 7,6), 5 mm MgCl2 , 10 mm Tris (pH 8), 0,05 mm EDTA (pH 8), 0,05 mm , 0,1% Triton X-100, 20% gliserol). Protein terkait dielusi dalam buffer Laemmli, dan interaksi protein–DNA ditentukan dengan western blotting menggunakan antibodi monoklonal tikus FLAG (Sigma-Aldrich, St. Louis, MO, AS). Urutan oligonukleotida berikut digunakan:
ETS 5′ACCGGAAGTACCGGAAGTACCGGAAGT 3′
AP1 5′ TGACTCATGAGTCAGTATGAGTCACAATGACTCACCT 3′
Uji angiogenesis ikan zebra
Percobaan dilakukan di tempat berlisensi untuk pengembangbiakan dan penggunaan hewan percobaan (Universitas Leiden) dan tunduk pada peraturan dan pedoman internal, yang menyatakan bahwa saran diambil dari badan kesejahteraan hewan untuk meminimalkan penderitaan bagi semua hewan percobaan yang ditempatkan di fasilitas tersebut. Pengujian ikan zebra yang dijelaskan tidak dianggap sebagai percobaan hewan berdasarkan Undang-Undang Percobaan pada Hewan (Wod, efektif 2014), undang-undang yang berlaku di Belanda sesuai dengan pedoman Eropa (arahan UE no. 2010/63/EU) tentang perlindungan hewan yang digunakan untuk tujuan ilmiah, karena larva yang tidak makan digunakan. Oleh karena itu, lisensi khusus untuk pengujian ini pada larva ikan zebra (< 5 hari) tidak diperlukan. Garis transgenik fli1a:gfp menghasilkan embrio di mana semua sel endotel ditandai oleh GFP. Ditambah dengan transparansi optiknya, ini memungkinkan visualisasi langsung angiogenesis. Selama 2 hari pertama perkembangan, pola berulang pembuluh darah intersegmental (ISV) dibentuk oleh tunas angiogenik dari sel endotel aorta dorsal yang tumbuh ke sisi dorsal batang tubuh tempat mereka saling terhubung untuk membentuk pembuluh anastomosis longitudinal dorsal (DLAV). Untuk percobaan yang dijelaskan di sini, embrio disimpan dalam air telur (60 μg·mL −1 garam laut; ~ 60 embrio/cawan) pada suhu 28 °C. Setelah pembuahan, perawatan obat ditambahkan langsung ke air telur. Setelah 24 jam, korion dikeluarkan secara mekanis menggunakan forsep. Embrio dibius menggunakan tricaine methanesulfonate (Sigma-Aldrich) pada konsentrasi akhir 0,003% dalam air telur, sekitar 10 menit sebelum pencitraan. Pencitraan pembuluh darah dilakukan dengan menggunakan mikroskop confocal Leica SP5 (Leica Microsystems, Wetzlar, Jerman) menggunakan lensa objektif ×10 atau ×20. Karakteristik pembuluh darah dari 20 embrio per kondisi dinilai.
Uji metatarsal
Embrio diisolasi dari uterus dan disimpan dalam PBS di atas es. Prosedur pembedahan metatarsal [ [ 54 ] ] dilakukan pada embrio tikus pada Hari ke-17 kehamilan. Metatarsal dikultur, dengan atau tanpa pengobatan obat, pada suhu 37 °C dalam cawan berlapis gelatin 0,1% dalam MEM Alpha Medium (Gibco) dan GlutaMAX (Gibco), yang mengandung 10% FBS (Gibco) dan penisilin-streptomisin (ICN Biomedicals, Aurora, OH, AS). Medium ini dilengkapi dengan 50 ng·mL −1 VEGF 165 manusia rekombinan (R&D Systems, Minneapolis, MN, AS). Medium kultur diganti setiap 2 hari. Setelah 5 hari kultur, pertumbuhan pembuluh dari metatarsal divisualisasikan dengan imunopewarnaan. Metatarsal dicuci dengan DPBS (Dulbecco’s PBS; Gibco) dan difiksasi dalam Zink Macrodex Formalin (PFA; Sigma-Aldrich) selama 15 menit. Pewarnaan metatarsal dilakukan menggunakan antibodi CD31 (BD Biosciences, San Diego, CA, AS) seperti yang dijelaskan sebelumnya [ [ 55 ] ]. Untuk analisis terperinci pembentukan pembuluh, konfigurasi pembuluh diubah menjadi gambar biner hitam-putih. Area vaskular (hitam) dalam piksel diukur menggunakan perangkat lunak komputer internal yang dikembangkan oleh Departemen Biologi Sel dan Kimia di LUMC. Dalam semua perawatan, total area vaskular telah dinormalisasi terhadap area yang meliputi tulang metatarsal [ [ 54 ] ]. Percobaan tikus disetujui oleh Komite Kelembagaan untuk Kesejahteraan Hewan dari Pusat Medis Universitas Leiden (LUMC) dan dilakukan sesuai dengan pedoman peraturan.
Kardiomiosit
Kardiomiosit yang berasal dari sel punca pluripoten yang diinduksi manusia (hiPSC-CM) (disediakan oleh R. Davis, LUMC) ditumbuhkan pada penutup kaca berukuran 5 mm dalam 100 μL Matrigel™ (Corning) dan 1 mL medium kultur [DMEM (Gibco) yang disuplemenkan dengan 10% FBS (Gibco), 450 μ m α-MTG, 0,05 mg·mL −1 l -asam askorbat 2-fosfat (Sigma-Aldrich), 2 mm GlutaMAX (Gibco), 0,5% Penisilin/streptomisin (ICN Biomedicals)]. Sel diinkubasi pada suhu 37 °C dengan atau tanpa senyawa yang diindikasikan (Gbr. 4D ). Setelah 16 jam pengobatan, sel difiksasi dengan 4% paraformaldehyde-phosphate buffered saline (PBS) selama 15 menit dan dipermeabilisasi dalam 0,2% Triton X-100-PBS selama 5 menit. Selanjutnya, sel dicuci dengan PBS 0,5% Tween dan diblokir dengan larutan yang mengandung PBS/0,5% Tween dan 5% BSA selama 30 menit. Imunostaining dilakukan dengan antibodi primer, Troponin I (Santa Cruz Biotechnology, Santa Cruz, CA, AS) dan anti-phospho-histone H2A.X (Ser139) (Cell Signaling Technology, Danvers, MA, AS), seperti yang dijelaskan sebelumnya [ [ 55 ] ]. Sel dicitrakan menggunakan mikroskop confocal Leica SP8.
Antibodi
Antibodi primer berikut digunakan: antibodi monoklonal tikus FLAG M2 (Sigma-Aldrich), antibodi monoklonal tikus anti-HA.11 (Covance, Princeton, NJ, AS), antibodi poliklonal kelinci anti-HA (Abcam, Cambridge, Inggris), antibodi anti-Troponin (Santa Cruz Biotechnology), antibodi anti-fosfo-histon H2A.X (Ser139) (Cell Signaling Technology). Antibodi sekunder berikut digunakan: antibodi anti-Rabbit Alexa Fluor® 488 dan antibodi anti-Mouse Alexa Fluor® 594 (Thermo Fisher Scientific).
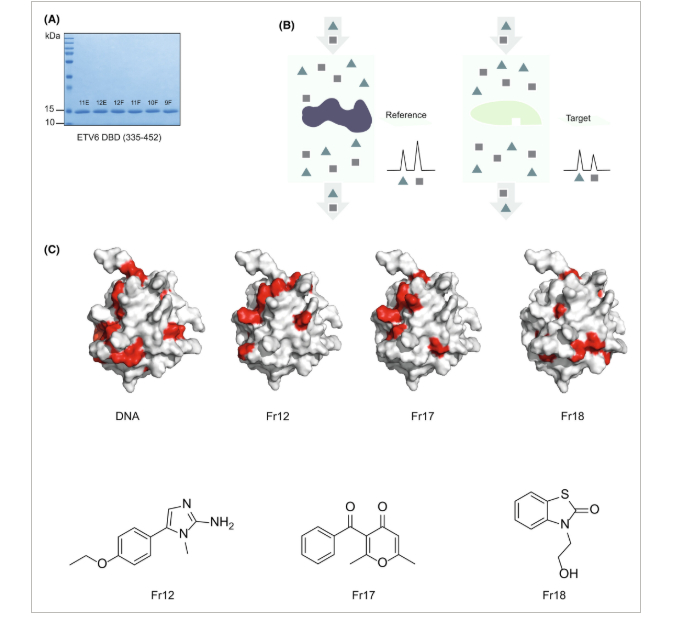
Validasi pengikatan fragmen ke EDBD
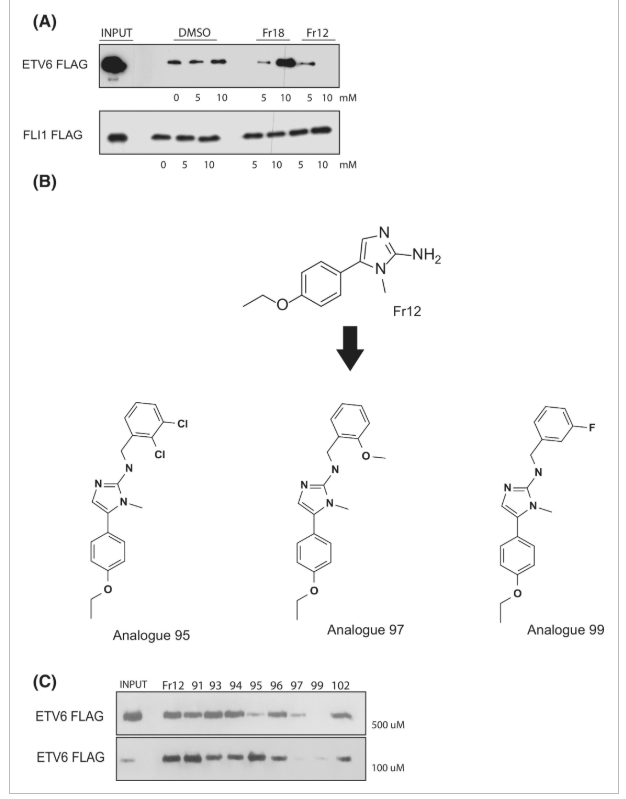
Untuk lebih mengkarakterisasi fragmen hit, kami membuat uji in vitro yang menggunakan situs pengikatan DNA ETS konsensus biotinilasi dan protein ETS yang diterjemahkan in vitro untuk merangkum pengikatan faktor ETS ke DNA. Uji yang dioptimalkan ini digunakan untuk menilai efek fragmen pada pengikatan protein ETS ke situs pengikatan DNA-nya. Gambar 2A mengilustrasikan setidaknya dua kelas hit yang berbeda: satu fragmen (Fr18) meningkatkan pengikatan DNA ETS, sedangkan fragmen yang berbeda (Fr12) secara signifikan menghambat pengikatan DNA ETS. Menariknya, fragmen tidak memiliki efek yang terdeteksi pada pengikatan faktor ETS terkait ke situs DNA konsensus (Gbr. 2A ), yang menunjukkan bahwa fragmen mungkin menunjukkan tingkat spesifisitas untuk targetnya. Afinitas fragmen yang sangat rendah untuk target (dalam kisaran mm ) tidak terduga karena ukurannya yang relatif kecil, dan meskipun ada bukti interaksi langsung antara fragmen hit dan EDBD, afinitasnya akan terlalu rendah untuk menimbulkan efek yang bermakna secara biologis. Untuk mengatasi masalah ini, kami menguji analog dari hit, yang memiliki struktur inti yang identik dengan hit fragmen asli, dan rantai samping kimia unik tambahan (Gbr. 2B ). Gambar 2C menunjukkan bahwa sebagian kecil analog fragmen menghambat pengikatan DNA ETS pada konsentrasi yang jauh lebih rendah (500–100 μm , peningkatan potensi 100 kali lipat), dibandingkan dengan hit fragmen, dalam uji pengikatan fungsional in vitro .
Secara kolektif, data ini menunjukkan bahwa penyaringan berbasis fragmen berpotensi mengidentifikasi perancah molekuler yang mengikat EDBD, dan pengoptimalan analog selanjutnya berpotensi menghasilkan senyawa dengan afinitas yang lebih unggul untuk protein target EDBD dibandingkan dengan fragmen asli. Pendekatan ini telah secara rutin diadopsi oleh industri sebagai uji bukti prinsip kesesuaian target untuk penyaringan skala yang lebih besar [ [ 46 ] ]. Dalam konteks ini, kami selanjutnya melakukan penyaringan berthroughput tinggi terhadap 400.000 senyawa.

Secara umum, dua jenis pemeriksaan molekul kecil dapat dilakukan untuk mengidentifikasi hit yang berpotensi menghambat aktivitas protein target tertentu: pemeriksaan pengikatan langsung komponen tunggal, seperti pemeriksaan TINS yang dijelaskan dalam Gambar 1 dan 2 ; dan pemeriksaan fungsional multikomponen di mana molekul kecil mengganggu proses biokimia tertentu, misalnya, interaksi protein-protein atau protein-DNA. Pengujian harus cepat, terukur, dan otomatis. Di sini, kami menerapkan pengujian fungsional untuk mengukur secara kuantitatif pengikatan EDBD ke situs pengikatan DNA konsensus. Untuk tujuan ini, kami menggunakan Homogeneous Time-Resolved Fluorescence (HTRF) [ [ 56 , 57 ] ] untuk menguji pengikatan EDBD berlabel epitop His 6 ke oligonukleotida terbiotinilasi yang mengandung tiga situs pengikatan ETS konsensus (lihat Gambar 3A ; Bagian bahan dan metode ). Singkatnya, sinyal FRET dihasilkan ketika fluorofor terkonjugasi dengan antibodi anti-His 6 (yang mengenali tag His dari EDBD) berada dalam jarak dekat dengan streptavidin XL-665 (yang mengenali bagian biotin dari oligonukleotida). Senyawa yang dapat mengganggu pengikatan protein ETS ke situs DNA menyebabkan hilangnya sinyal fluoresensi (Gbr. 3A ). Dengan menggunakan uji ini, 400.000 molekul kecil disaring dalam uji primer. Validasi lebih lanjut untuk menilai reproduktifitas (lihat bagian Bahan dan metode ), ditambah dengan analisis spektrometri massa kromatografi cair (LCMS) dari senyawa tersebut, menghasilkan pemilihan empat hit, yang secara terukur menghambat pengikatan DNA EDBD, untuk penyederhanaan diberi nama senyawa A–D. Struktur kimia kandidat hit digambarkan dalam Gbr. 3B .
Validasi biokimia senyawa hit dalam uji pengikatan fungsional in vitro
Gambar 3C menunjukkan bahwa senyawa hit secara efisien menghambat pengikatan faktor ETS ke situs pengikatan konsensus DNA-nya tetapi gagal menghambat pengikatan DNA JUN ke situs DNA konsensus yang terkait tetapi berbeda, konsisten dengan gagasan bahwa penghambatan tersebut bersifat spesifik terhadap pengikatan DNA EDBD. Selain ETV6, senyawa hit juga menghambat pengikatan DNA faktor ETS lainnya, termasuk ERG dan FLI1, yang menunjukkan bahwa penghambatan tersebut tidak terbatas pada faktor transkripsi ETS tertentu, yang mungkin mencerminkan tingkat kesamaan struktural EDBD secara keseluruhan yang tinggi dalam keluarga faktor transkripsi ETS.
Senyawa hit secara signifikan menghambat proliferasi sel tumor
Karena senyawa hit menghambat pengikatan ke DNA dari beberapa faktor transkripsi ETS yang berbeda, selanjutnya kami menguji efek senyawa pada proliferasi lini sel tumor. Beberapa lini sel kanker (melanoma, karsinoma pankreas, karsinoma prostat dan sarkoma Ewing), yang fungsi ETS diketahui rusak, diuji, dan diperoleh hasil yang sebanding. Gambar 3D menyajikan data representatif pada dua lini sel, yang mengandung translokasi kromosom yang mengakibatkan fungsi FLI1 atau ERG yang tidak normal pada sel sarkoma Ewing [ [ 58 ] ] dan karsinoma prostat [ [ 59 , 60 ] ], masing-masing. Proliferasi lini sel kanker ditekan secara signifikan oleh senyawa A, B dan D, masing-masing menunjukkan nilai konsentrasi penghambatan setengah maksimal (IC50) dalam kisaran 1–10 μ m (Gbr. 3D ). Sebaliknya, senyawa tersebut tidak secara signifikan memblokir proliferasi HUVEC primer normal pada tingkat yang sama dengan sel tumor.
Senyawa hit menghambat perkecambahan angiogenik
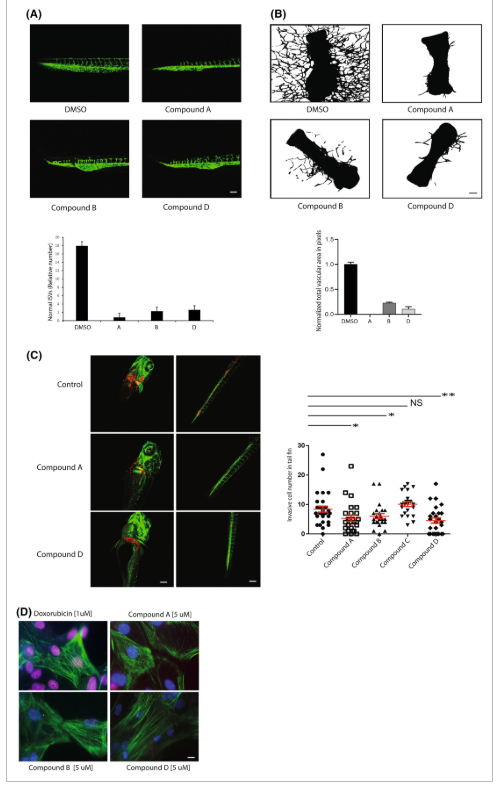
Bahasa Indonesia: Selain mengendalikan proliferasi sel tumor secara langsung, faktor ETS juga memainkan beberapa peran berbeda lainnya dalam biologi stroma tumor, terutama dalam vaskularitas tumor, karena faktor ETS sangat diperlukan untuk perluasan pembuluh darah melalui proses angiogenesis [ [ 31 – 33 ] ]. Dalam hal ini, kami menguji efek senyawa dalam tiga uji angiogenesis yang berbeda. Pertama, kami menggunakan garis zebrafish transgenik fli1 a:gfp yang menghasilkan embrio yang semua pembuluh darahnya ditandai oleh GFP, yang, ditambah dengan sifat embrio yang transparan secara optik, memungkinkan visualisasi angiogenesis yang sistematis dan dinamis. Gambar 4A menunjukkan bahwa inkubasi embrio zebrafish dengan 10 μm senyawa A, B dan D selama 16 jam mengakibatkan gangguan angiogenesis yang jelas, yang dimanifestasikan oleh pengurangan jumlah pembuluh, lintasan pembuluh yang tidak normal dan penghentian prematur tunas aorta dorsal (Gbr. 4A ). Efeknya paling jelas terlihat pada senyawa A, B, dan D (Gbr. 4A ), sementara inkubasi dengan obat C tidak mengganggu pembentukan pembuluh darah secara signifikan. Efek ini tidak terkait dengan toksisitas umum yang nyata.
Bahasa Indonesia: Untuk lebih mengkarakterisasi efek molekul kecil pada angiogenesis, kami menggunakan uji metatarsal tikus fetus ex vivo , yang merangkum banyak fitur penting angiogenesis yang diamati secara in vivo [ [ 54 ] ]. Metatarsal diisolasi dari fetus pada tahap embrio E17.5 dan diinkubasi dalam medium tertentu yang mengandung faktor pertumbuhan endotel vaskular (VEGF) dengan atau tanpa 1 μ m senyawa tersebut. Pembentukan pembuluh dipantau dengan mikroskop cahaya fase kontras sejak Hari ke-2. Senyawa A, B, dan D masing-masing menghambat angiogenesis ektopik pada metatarsal yang dikultur. Pewarnaan imunofluoresensi dengan molekul adhesi sel endotel dan trombosit 1 (PECAM-1) (antibodi CD31) dilakukan untuk memvisualisasikan pertumbuhan pembuluh metatarsal. Gambar konfokal pembuluh mikro ditunjukkan (Gbr. 4B ). Analisis kuantitatif percabangan pembuluh dilakukan berdasarkan jumlah piksel di area pembuluh (Gbr. 4B ). Gambar 3D menunjukkan bahwa kandidat yang terkena HTS secara efektif menghambat proliferasi sel tumor; namun, senyawa tersebut tidak secara signifikan memblokir proliferasi sel endotel, yang mengekspresikan kadar aktivitas protein ETS yang normal, pada tingkat yang sama dengan sel tumor, konsisten dengan gagasan bahwa efek senyawa yang diamati pada angiogenesis terutama dihasilkan dari penghambatan perkecambahan endotel dibandingkan dengan proliferasi sel endotel.
Mengingat pentingnya faktor ETS dalam proliferasi dan pergerakan sel, ditambah dengan peran mereka dalam angiogenesis sprouting, inhibitor ETS dapat menekan invasi dan metastasis sel tumor. Di sini, kami menggunakan uji xenotransplantasi zebrafish Tg ( fli1 :gfp) untuk menyelidiki ekstravasasi dan intravasasi sel. Sekitar 400 sel kanker payudara manusia MDA-MB-231 (diberi label genetik dengan mCherry) disuntikkan ke dalam saluran Cuvier 48 jam setelah pembuahan. Embrio zebrafish yang disuntikkan diperlakukan dengan senyawa dengan penambahan air kultur. Gambar 4C menunjukkan bahwa penyebaran sel kanker payudara ke arah kepala dan ekor berkurang secara signifikan dengan adanya senyawa A, B dan D (10 μ m ).
Secara keseluruhan, data ini menunjukkan bahwa, selain secara langsung menghambat proliferasi sel tumor, senyawa sasaran terpilih memberikan efek penghambatan pada pertumbuhan angiogenik dan intravasasi sel tumor.
Evaluasi senyawa hit kardiotoksisitas
Toksisitas jangka pendek dan jangka panjang merupakan masalah mendasar yang terkait dengan perawatan kemoterapi, khususnya perawatan yang merusak DNA. Misalnya, doksorubisin, reagen kemoterapi yang banyak digunakan, mengaktifkan mesin respons kerusakan DNA yang pada akhirnya mengakibatkan apoptosis dan kardiotoksisitas jangka panjang terkait, yang menyebabkan kerusakan jaringan jantung dan potensi gagal jantung [ [ 61 ] ]. Oleh karena itu, kardiotoksisitas adalah salah satu pertimbangan terpenting saat mengembangkan terapi baru dengan tujuan meningkatkan perawatan saat ini. Di sini, kami menggunakan kardiomiosit yang berasal dari sel punca pluripoten yang diinduksi manusia (hiPSC-CM) untuk mengevaluasi kardiotoksisitas senyawa hit, memanfaatkan antibodi yang menargetkan histon H2AX terfosforilasi. Gambar 4D menunjukkan bahwa senyawa hit menunjukkan tingkat kerusakan DNA yang relatif minimal jika dibandingkan dengan obat kemoterapi konvensional, doksorubisin.
Diskusi
Bahasa Indonesia: Sementara dua dekade terakhir menyaksikan upaya yang cukup besar untuk menargetkan reseptor dan kinase untuk menghambat pertumbuhan tumor [ [ 1 – 5 , 11 – 17 ] ], keberhasilan klinis dari terapi tersebut sering terhambat oleh resistensi dan toksisitas obat [ [ 6 – 10 ] ]. Meskipun penargetan faktor transkripsi secara historis diabaikan karena kompleksitas struktural dan daerah yang secara intrinsik tidak teratur, kemajuan terkini dalam penemuan obat dan kimia modern telah menyalakan kembali minat dalam menargetkan protein tersebut. Keluarga faktor transkripsi ETS sering rusak dalam kanker [ [ 26 ] ], dan mereka terletak di hilir jalur pensinyalan utama [ [ 62 , 63 ] ], yang salah diatur selama tumourigenesis. Dengan demikian, molekul yang secara spesifik menghambat fungsi ETS yang tidak normal berpotensi membatasi toksisitas nyata dan resistensi terapi yang didapat.
Pendekatan untuk mengidentifikasi inhibitor aktivitas faktor ETS spesifik [ [ 36 , 38 , 41 ] ] atau protein fusi ETS [ [ 37 ] ] telah mencakup metode kimia komputasional dan penyaringan fungsional, yang telah menghasilkan molekul kandidat yang menargetkan situs pengikatan DNA konsensus ETS [ [ 38 ] ], mesin biogenesis ribosomal [ [ 41 ] ] atau interaksi dengan helicase RNA [ [ 37 ] ]. Untuk mengeksplorasi potensi ligandabilitas faktor ETS dan untuk mengidentifikasi senyawa, yang secara langsung dapat memblokir fungsi faktor ETS melalui penghambatan pengikatan ETS ke situs pengikatan DNA konsensusnya, kami telah melakukan penyaringan berbasis molekul kecil/fragmen bukti prinsip. Dua jenis penyaringan telah dilakukan: (a) penyaringan berbasis fragmen, menggunakan teknologi TINS, yang memilih perancah molekuler yang secara langsung mengikat EDBD; (b) HTS molekul kecil mirip timbal untuk mengidentifikasi senyawa yang mampu memblokir pengikatan EDBD ke situs DNA konsensusnya. Skrining berbasis fragmen menunjukkan bahwa perancah molekuler kecil dapat berinteraksi dengan EDBD (Gbr. 1 dan 2 ). Afinitas fragmen yang relatif lemah untuk target tidak cukup untuk merusak fungsi faktor ETS pada dosis yang bermakna secara terapeutik. Namun, keuntungan dari perpustakaan fragmen adalah keragaman kimianya dan berat molekul fragmen yang relatif rendah, yang secara signifikan meningkatkan peluang mengidentifikasi hit primer yang dapat berevolusi menjadi senyawa timbal melalui hubungan struktur-aktivitas, yang dapat menunjukkan karakteristik yang menguntungkan secara farmakologis [ [ 45 , 46 ] ]. Memang, kami menunjukkan bahwa analog hit fragmen yang lebih kompleks secara kimia dan lebih besar dapat menghambat pengikatan DNA EDBD pada konsentrasi dalam kisaran 100 kali lipat lebih rendah daripada kandidat hit primer. Keberhasilan relatif dari pengukuran interaksi langsung antara target dan sasaran menunjukkan bahwa pendekatan yang sebanding dapat menghasilkan inhibitor ETS, seperti pustaka molekul kecil yang dikodekan DNA, yang memungkinkan penyaringan dalam skala besar (hingga miliaran senyawa), atau penyaringan pustaka yang lebih kecil seperti timbal. Kemudahan pemurnian EDBD yang relatif mudah ditambah dengan kecepatan prosedur penyaringan dapat menjadikan ini pilihan yang menarik [ [ 64 ] ].
Pendekatan alternatif untuk penyaringan ‘pengikat langsung’ adalah penyaringan fungsional, dalam hal ini, memblokir pengikatan spesifik EDBD ke situs DNA konsensus. Pengujian HTRF cepat dan dapat diskalakan untuk menyaring perpustakaan molekul kecil (> 400.000 molekul). Dalam penelitian kami, pendekatan seperti itu menghasilkan senyawa, yang dapat mengganggu pengikatan EDBD ke DNA dan menghambat proliferasi sel tumor (yang mengandung protein ETS yang diaktifkan secara tidak sah). Faktor ETS seperti ETV6, ETS1, FLI1 dan ERG sangat penting untuk menumbuhkan angiogenesis [ [ 31 – 33 ] ], salah satu ciri khas kanker, dan kami menemukan bahwa senyawa yang dipilih dapat secara efisien menghambat proses ini, meningkatkan kemungkinan bahwa penghambatan ETS pada akhirnya dapat mewakili pendekatan membunuh dua burung dengan satu batu untuk penargetan tumor. Senyawa tersebut tidak secara signifikan merusak pengikatan JUN ke situs DNA konsensusnya, konsisten dengan gagasan bahwa senyawa tersebut menunjukkan spesifisitas. Namun, mereka tidak selektif dalam menghambat faktor ETS tertentu dan malah memblokir pengikatan DNA dari beberapa anggota keluarga ETS yang berbeda (Gbr. 3C ), yang dapat mencerminkan konservasi urutan asam amino keseluruhan yang tinggi dari EDBD dan mode pengikatan DNA yang sebanding. Perlu dicatat, senyawa yang diidentifikasi dalam dua penyaringan yang berbeda tidak memiliki kesamaan struktural yang jelas, yang tidak mengherankan karena pustaka senyawa yang digunakan (pustaka fragmen dan pustaka molekul kecil dengan massa molekul yang lebih tinggi) jelas berbeda. Hal ini juga dapat mencerminkan fakta bahwa penyaringan tersebut juga berbeda secara fungsional: penyaringan pustaka fragmen mengidentifikasi molekul, yang berpotensi mengikat langsung ke target ‘bebas’ (tidak terikat) sementara penyaringan molekul kecil multikomponen mengidentifikasi senyawa yang berpotensi mengganggu pengikatan EDBD/DNA. Dengan demikian, konformasi EDBD dapat berbeda dalam setiap kasus dan tidak dapat dikesampingkan bahwa senyawa dalam penyaringan terakhir berinteraksi dengan kompleks EDBD/DNA. Memang, kurangnya spesifisitas EDBD yang jelas dan potensinya yang relatif tinggi dalam pengujian berbasis sel akan konsisten dengan pandangan ini. Inhibitor ETS generik dapat bernilai karena mereka cenderung menghambat proliferasi sel tumor secara lebih efisien; namun, bukan tidak mungkin bahwa, secara in vivo , inhibitor tersebut mungkin menunjukkan efek samping toksik, karena peran mereka dalam jaringan pensinyalan sel normal. Terkait dengan ini, bagaimanapun, perlu dicatat, bahwa senyawa tersebut tidak dikaitkan dengan tingkat kerusakan DNA yang tinggi pada kardiomiosit yang dikultur dibandingkan dengan obat kemoterapi yang umum digunakan, doksorubisin (Gbr. 4D ). Selain itu, embrio ikan zebra menoleransi jumlah senyawa μm yang rendah (Gbr. 4 ) .
Apakah ada solusi yang memungkinkan untuk mengatasi dua tantangan utama dalam menargetkan anggota keluarga ETS, yaitu, identifikasi inhibitor spesifik, yang secara eksklusif menargetkan satu anggota keluarga, dan mengatasi masalah afinitas rendah senyawa untuk targetnya? Sejumlah pendekatan dapat diambil. Pertama, meskipun ada tantangan teknis yang signifikan untuk diatasi, penyaringan pustaka terhadap protein ETS murni, fungsional, dan panjang penuh, alih-alih EDBD yang terisolasi, dapat meningkatkan spesifisitas, terutama jika senyawa yang ditabrak disaring balik terhadap anggota keluarga ETS murni lainnya. Kedua, dan digabungkan dengan ini, kemajuan signifikan dalam konstruksi pustaka kovalen, keragaman, dan metode penyaringan [ [ 65 ] ] dapat sangat meningkatkan afinitas senyawa karena mereka mengikat secara ireversibel ke targetnya.