
Abstrak
Antagonis protein morfogenetik tulang (BMP) semakin dikaitkan dengan perkembangan kanker kolorektal (CRC). Pensinyalan BMP beroperasi secara berlawanan dengan jalur pensinyalan WNT, yang menopang pemeliharaan sel punca dan pembaruan diri epitel usus normal. BMP yang berkurang dan pensinyalan WNT yang meningkat menyebabkan perluasan kompartemen sel punca dan hiperproliferasi sel epitel, karakteristik penentu CRC. Chordin-like-2 (CHRDL2) adalah antagonis BMP yang disekresikan, dengan ekspresi berlebih dikaitkan dengan prognosis yang buruk dan varian pada gen yang terbukti terkait dengan peningkatan risiko CRC. Namun, mekanisme terperinci yang berkontribusi pada CRC tidak diketahui. Dalam penelitian ini, kami mengeksplorasi dampak ekspresi berlebih CHRDL2 pada sel CRC untuk menyelidiki apakah penghambatan CHRDL2 terhadap pensinyalan BMP mengintensifkan pensinyalan WNT dan meningkatkan fenotipe sel punca kanker dan respons terhadap pengobatan. Pendekatan penelitian kami menggabungkan lini sel kanker 2D yang direkayasa untuk mengekspresikan CHRDL2 secara berlebihan dan model organoid 3D yang diobati dengan CHRDL2 ekstrinsik, dilengkapi dengan analisis sekuensing RNA. CHRDL2 ditemukan dapat meningkatkan kelangsungan hidup organoid dan sel CRC selama kemoterapi dan pengobatan iradiasi karena aktivasi jalur respons kerusakan DNA. Organoid yang diobati dengan CHRDL2 yang disekresikan menunjukkan peningkatan kadar penanda sel punca dan diferensiasi yang berkurang, sebagaimana dibuktikan oleh berkurangnya tunas vili. Analisis RNA-seq mengungkapkan bahwa CHRDL2 meningkatkan ekspresi penanda sel punca, pensinyalan WNT, dan jalur terkait kanker lain yang mapan melalui penghambatan BMP. Temuan-temuan ini secara kolektif menunjukkan bahwa ekspresi berlebihan CHRDL2 dapat memengaruhi respons terhadap terapi CRC dengan meningkatkan perbaikan DNA dan potensi sel punca sel kanker, dan perannya sebagai biomarker harus dieksplorasi lebih lanjut.
Singkatan
ATM
Ataksia-telangiektasia bermutasi
BMI1
Homolog daerah penyisipan 1 Mo-MLV limfoma B
BMP
protein pembentuk tulang
CHRDL2
seperti Chordin 2
CRC
kanker kolorektal
Bahasa Indonesia: CSC
sel induk kanker
Bahasa Inggris DSB
pemutusan untai ganda
Bahasa Indonesia: ISC
sel induk usus
LGR5
reseptor 5 yang mengandung pengulangan kaya leusin yang digabungkan dengan protein G
LGR6
reseptor leusin kaya pengulangan yang mengandung protein G yang digabungkan 6
NTB
Jalur pensinyalan WNT
γH2AX
Anggota keluarga histon H2A X
1 Pendahuluan
Kanker kolorektal (CRC) menempati peringkat ketiga kanker dengan prevalensi tertinggi di dunia, dengan lebih dari 1 juta kasus yang dilaporkan pada tahun 2020 menurut GLOBOCAN [ [ 1 ] ]. Berasal dari mutasi dalam sel epitel usus, CRC menyebabkan pembentukan polip, adenokarsinoma, dan akhirnya kanker metastasis. Meskipun banyak jalur pensinyalan terganggu pada CRC, jalur pensinyalan WNT/β-catenin adalah yang paling sering terpengaruh, terlalu aktif pada hampir semua kasus CRC [ [ 2 ] ].
Sinyal WNT adalah jalur fundamental yang mengatur proliferasi dan nasib sel induk intestinal (ISC). Aktivasi WNT terlokalisasi di dasar kripta intestinal, tempat ia mengendalikan nasib dan pembaruan ISC, komponen vital dari pemeliharaan intestinal [ [ 3 ] ]. ISC kripta terorganisasi secara hierarkis, dengan ISC yang berproliferasi cepat berada di dasar kripta dan ISC yang berproliferasi lebih lambat atau regeneratif sedikit bergeser dari dasar kripta pada posisi +4 [ [ 4 , 5 ] ]. Subset ketiga sel yang berproliferasi cepat yang lebih matang bermigrasi lebih jauh ke atas kripta ke zona transit amplifying (TA). ISC diidentifikasi dengan ekspresi penanda LGR5+, dan sel siklus lambat pada posisi +4 sebelumnya telah diidentifikasi dengan adanya penanda BMI1+, serta HPOX dan TERT, tetapi dengan data yang saling bertentangan tentang penanda spesifik dan peran sel-sel ini [ [ 6 ] ].
Pensinyalan WNT bertransduksi dengan mengisolasi kompleks destruksi β-catenin, meningkatkan kadar β-catenin intraseluler, dan mengaktifkan jalur sel punca dan onkogenik dalam kripta sel punca [ [ 7 , 8 ] ]. Gangguan pensinyalan WNT menghilangkan fenotip progenitor dalam kripta dan menyebabkan hilangnya kripta [ [ 9 ] ]. Meskipun pensinyalan WNT diperlukan untuk mempertahankan lapisan usus dan pembentukan kripta, pensinyalan WNT yang berkelanjutan atau meningkat dapat menyebabkan hiperproliferasi dan transformasi onkogenik dalam ISC [ [ 10 ] ]. Pensinyalan WNT telah terbukti bekerja dalam gradien berlawanan dengan pensinyalan BMP, yang ditemukan di vili usus dan meningkatkan diferensiasi dan pematangan sel [ [ 10 – 12 ] ]. Gradien pensinyalan BMP dan WNT ini merupakan faktor pengendali utama dalam arsitektur kripta-vili dan homeostasis usus.
Berbeda dengan pensinyalan WNT, pensinyalan BMP terlokalisasi di kompartemen terdiferensiasi kripta/vili dan membantu diferensiasi, proliferasi, dan migrasi seluler [ [ 13 ] ]. BMP telah terbukti memiliki efek paradoks pada kanker, dengan ligan spesifik yang bekerja untuk menghambat dan mendorong tumorigenesis di berbagai jaringan dan konteks [ [ 14 – 17 ] ]. BMP termasuk dalam superfamili TGF-β dan mengikat kompleks reseptor kinase serin-treonin transmembran I dan II (BMPR I dan II) [ [ 18 ] ]. Hal ini memulai fosforilasi reseptor tipe I oleh reseptor tipe II, memicu fosforilasi SMAD terkait reseptor yang kemudian membentuk kompleks dengan SMAD4 dan bertranslokasi ke nukleus untuk mengatur transkripsi gen [ [ 19 ] ]. Sementara sel epitel dan mesenkim mengekspresikan BMP dan reseptornya, antagonis BMP terutama ditemukan di mesenkim. Di dalam usus, mereka sebagian besar diekspresikan oleh miofibroblast kriptal usus dan sel otot polos. Antagonis ini memblokir sinyal BMP di kompartemen sel induk, mempertahankan tingkat sinyal WNT yang tinggi dan dengan demikian sel induk [ [ 11 ] ].
Antagonis BMP dapat mengikat langsung ke BMP atau reseptornya [ [ 20 ] ]. Beberapa antagonis BMP yang telah diteliti dengan baik meliputi Noggin, yang telah terlibat dalam mempromosikan tumorigenesis kanker kulit dan payudara, dan Gremlin (GREM1 dan 2), dengan represi GREM1 yang terbukti menghambat proliferasi sel tumor. Keluarga protein Chordin juga telah terlibat dalam CRC, termasuk Chordin, Chordin-like 1 (CHRDL1) dan Chordin-like 2 (CHRDL2) [ [ 21 – 24 ] ]. Salah satu antagonis BMP yang paling banyak dipelajari, Noggin, sebelumnya telah terbukti menghambat pensinyalan BMP dalam model tikus, yang mengakibatkan pembentukan banyak kripta ektopik yang tegak lurus dengan sumbu kripta-vilus [ [ 12 ] ]. Demikian pula, ekspresi berlebihan GREM1 pada sindrom poliposis campuran herediter (HMPS) menyebabkan persistensi atau perolehan kembali sifat sel punca pada sel negatif LGR5 di luar ceruk sel punca. Kripta ektopik, peningkatan proliferasi dan neoplasia usus [ [ 25 ] ]. Secara bersamaan, hal ini menunjukkan bahwa penghapusan sinyal BMP melalui antagonisnya menyebabkan pembentukan kualitas seperti sel punca pada sel epitel usus, yang mengarah pada transformasi onkogenik.
CHRDL2 adalah antagonis BMP yang mencegah ligan BMP 2, 4 dan 6 berinteraksi dengan reseptor permukaan sel yang sesuai [ [ 22 , 26 ] ]. CHRDL2 telah terbukti mengikat langsung ke BMP, mencegah pensinyalan melalui fosfo SMAD1/5. Lebih jauh lagi, CHRDL2 menghambat efek pensinyalan BMP pada penghambatan proliferasi dan apoptosis [ [ 27 ] ]. Peningkatan mRNA CHRDL2 telah diamati pada kanker usus besar, payudara, hati dan prostat [ [ 27 , 28 ] ] dan kadar yang tinggi memprediksi prognosis yang buruk dan berkorelasi dengan peningkatan ukuran tumor dan stadium TNM selanjutnya [ [ 27 ] ]. CHRDL2 telah disorot sebagai biomarker protein sirkulasi potensial untuk CRC, di mana kadar CHRDL2 yang lebih tinggi yang diprediksi secara genetik dikaitkan dengan peningkatan risiko CRC [ [ 29 ] ]. Peran fungsional CHRDL2 yang tepat dalam kanker ini tidak selalu jelas, tetapi telah terbukti meningkatkan proliferasi sel, migrasi, dan invasi dalam lini sel osteosarkoma melalui regulasi jalur PI3k/AKT melalui pengikatan ke BMP9 [ [ 30 ] ]. Namun, peran sinyal BMP dalam kanker, dan oleh karena itu, efek penghambatan BMP oleh CHRDL2 dalam kanker masih kurang dipahami.
Dalam penelitian ini, kami menggunakan lini sel CRC yang direkayasa untuk mengekspresikan CHRDL2 secara berlebihan secara stabil dengan cara yang dapat diinduksi, untuk menyelidiki jalur seluler dan transkripsi yang diaktifkan oleh ekspresi CHRDL2 dan penghambatan BMP. Kami telah menunjukkan bahwa CHRDL2 memiliki efek yang terukur pada proliferasi sel dan secara signifikan mengubah respons terhadap kemoterapi yang merusak DNA. Untuk mendapatkan wawasan yang lebih mendalam tentang peran CHRDL2 dalam pemeliharaan dan diferensiasi sel punca, kami membudidayakan organoid usus 3D yang dilengkapi dengan bentuk CHRDL2 yang disekresikan. Secara kolektif, temuan kami menunjukkan bahwa CHRDL2 memodulasi jalur sel punca dalam CRC, yang berpotensi memengaruhi respons terhadap intervensi kemoterapi umum.
2 Bahan dan Metode
2.1 Kultur sel dan pemeliharaannya
Lini sel adenokarsinoma kolorektal manusia yang diabadikan Caco-2 (RRID:CVCL_0025), COLO 320 (RRID:CVCL_1989), LS180 (RRID:CVCL_0397) dan RKO (RRID:CVCL_0504) diperoleh dari ATCC dan diautentikasi dengan analisis STR (Eurofins, Ebersberg, Jerman) dalam waktu 3 tahun penggunaan untuk penelitian ini. Morfologi lini sel terus diperiksa untuk memastikan keaslian. Semua percobaan dilakukan dengan sel yang bebas mikoplasma. Sel dipelihara dalam medium Eagle yang dimodifikasi Gibco Dulbecco (DMEM) (Merck, Gillingham, Inggris) yang dilengkapi dengan 10% serum sapi janin (FBS) (Merck), dan 1% penisilin-streptomisin (Merck). Sel tumbuh dalam atmosfer yang dilembabkan pada suhu 37 °C dengan 5% CO 2 . Subkultur dilakukan setiap 72 jam untuk mempertahankan konfluensi sel <80%. 2.2 Pembuatan dan validasi lini sel yang mengekspresikan CHRDL2 secara berlebihan cDNA CHRDL2 dengan panjang penuh (Genecopoeia GC-H1938, Caltag Buckingham, Inggris) dikloning ke dalam pCW57.1 (Addgene #41393, Watertown, MA, AS) menggunakan teknologi Gateway (Invitrogen, Thermo Fisher, Waltham, MA, AS), diikuti dengan validasi melalui sekuensing Sanger dan pencernaan restriksi. Vektor tersebut kemudian ditransfeksi ke dalam sel HEK293 bersama dengan vektor pengemasan virus (sistem generasi ke-2—pCMV-dR8.2 dan pCMV-VSV-G) menggunakan Lipofectamine 2000 (Invitrogen, Fisher Scientific, Loughborough, Inggris). Media yang mengandung virus dikumpulkan, disterilkan, dan titer diukur (Go-Stix, Takara Bio, London, Inggris). Lini sel CACO2, COLO320, LS180, dan RKO ditransduksi, dan sel dengan pCW57.1-CHRDL2 terintegrasi dipilih dengan puromisin. Untuk mengonfirmasikan ekspresi berlebih, doksisiklin ditambahkan pada (0,1 μg·mL −1 , 1 μg·mL −1 (CHRDL2+) atau 10 μg·mL −1 (CHRDL2++) RNA diekstraksi (RNeasy, QIAGEN, Manchester, Inggris) dan diukur dengan reaksi berantai polimerase transkriptase balik waktu nyata (qPCR) menggunakan teknologi TaqMan (Hs00248808_m1) sesuai dengan protokol pabrikan (Applied Biosystems, Fisher Scientific, Loughborough, Inggris). Setiap pengujian diulang sebanyak tiga kali. 2.3 Western blot Untuk deteksi protein intraseluler, sel dilisiskan dengan resuspensi dalam buffer RIPA. Untuk ekspresi protein yang disekresikan, sel yang diberi ekspresi doksisiklin pada 10 mg·mL −1 diinkubasi selama 72 jam. Media dikumpulkan dan dipekatkan melalui filter sentrifugal Amicon ® Ultra (Merck) dengan ukuran pori 30 kDa. Media kemudian diencerkan 1:25, 1:25 dan 1:100 dengan buffer RIPA. Untuk protein intraseluler, 30 μg protein dimuat per sampel. Sampel protein dipisahkan melalui elektroforesis gel poliakrilamid natrium dodecyl sulfate 4–12% dalam kondisi denaturasi dan kemudian dipindahkan ke membran nitrocellulose (Millipore, Merck, Gillingham, UK) di bawah 20 V. Membran diblokir dengan susu 5% selama 1 jam pada suhu kamar. Membran kemudian diinkubasi dengan antibodi primer dalam TBST-5% BSA semalaman pada suhu 4 °C. Membran kemudian dicuci dengan TBST. Antibodi sekunder ditambahkan selama 1 jam pada suhu kamar. Membran dicitrakan melalui inkubasi dengan Enhanced chemiluminescence (ECL). Rasio kerapatan optik pita diukur dengan sistem analisis citra gel (Bio-Rad, Watford, Inggris) dan dinormalisasi ke B-aktin sebagai kontrol pemuatan. Untuk deteksi protein ekstraseluler, pewarnaan ponceau dikuantifikasi dan digunakan sebagai kontrol pemuatan. Antibodi berikut digunakan: CHRDL2 (1:1000, Nomor katalog: AF2448, Bio-techne, Abingdon, Inggris), P-SMAD1/5 (1:1000. Nomor katalog: #9516, Cell Signalling, Danvers, MA, AS), Goat anti-mouse (1:2000. Nomor katalog: ab205719, Abcam, Cambridge, Inggris), Goat anti-rabbit (1:5000. Nomor katalog: p0448, Dako, Agilent, Stockport, Inggris). 2.4 Uji proliferasi sel Untuk menilai proliferasi sel selama ekspresi berlebihan CHRDL2, sel-sel ditanam pada kepadatan 5 × 103 sel per sumur dalam pelat 96 sumur dengan 8 replikasi per kondisi: kontrol DMSO, perlakuan CHRDL2+ atau CHRDL2++. Proliferasi sel dinilai melalui uji MTS (CellTiter 96 ® Promega, Southampton, Inggris) pada 24, 48, dan 72 jam. 2.5 Aliran sitometri Sel COLO320 ditanam dalam pelat enam sumur dengan kepadatan 1 × 105 sel /sumur dalam kondisi media standar, dilengkapi dengan perlakuan DMSO atau CHRDL2++. Untuk analisis sitometri aliran kemoterapi, sel diperlakukan dengan 25 μ m Oxaliplatin. Sel ditumbuhkan selama 48 jam sebelum dipanen dengan tripsinisasi dan dicuci sekali dengan PBS dingin. Untuk menyelidiki perkembangan siklus sel, sel disuspensikan kembali dalam kontrol PBS atau Hoechst 33 342 (62 249; Thermo Scientific, Fisher Scientific, Loughborough, Inggris) yang diencerkan dalam PBS dan diinkubasi pada suhu 37 °C selama 2 jam dengan sedikit agitasi. Akhirnya, sampel dipelet dan disuspensikan kembali dalam PBS, dan analisis sitometri aliran dilakukan menggunakan sistem ACEA Novocyte (Agilent). Persentase sel dalam fase siklus sel dianalisis melalui perangkat lunak Novocyte. Untuk menyelidiki proporsi sel Ki67+, sel diinkubasi dengan Ki67 BV711 (407-5698-80; Thermo Scientific) selama 20 menit sebelum analisis. Untuk menyelidiki apoptosis, sel diwarnai dengan Zombie Aqua (423 101; BioLegend, London UK) dan antibodi annexin-V (V13242, Thermo Scientific) masing-masing selama 30 menit pada suhu ruangan. Selanjutnya, sel dicuci sekali dengan PBS dan dianalisis dengan flow cytometry menggunakan perangkat lunak pabrikan. 2.6 Uji pembentukan koloni Untuk menilai kemampuan sel tunggal untuk menghasilkan koloni dan kemampuan bertahan hidup sel, uji klonogenik dilakukan. Sel ditanam pada 100 sel per sumur dari pelat 6 sumur. Sel diobati dengan pengobatan doksisiklin seperti sebelumnya dalam berbagai konsentrasi 10 μg·mL −1 , 1 μg·mL −1 dan 0,1 μg·mL −1 atau DMSO. Pelat diinkubasi selama 2 minggu sampai koloni yang terlihat terbentuk. Setiap 72 jam, pengobatan doksisiklin dan DMSO disegarkan. Setelah 2 minggu, sel difiksasi dengan >98% metanol pada −20 °C dan diwarnai dengan pewarna kristal violet (0,5%, dalam 20% metanol). Koloni dihitung melalui Imag J.
2.7 Uji dosis-respons obat
Untuk menilai kemampuan lini sel untuk menahan pengobatan dari obat kemoterapi komersial, kurva dosis-respons obat dilakukan. Obat kemoterapi yang digunakan adalah sebagai berikut: 5-fluorouracil (5FU), irinotecan dan oxaliplatin (Merck). Pengenceran serial dilakukan dalam media kultur sel standar untuk memberikan berbagai konsentrasi (5FU 0–10.000 μ m , oxaliplatin 0–4.000 μ m , irinotecan 0–500 μ m ). Sel dari kultur disemai pada pelat 96-sumur pada kepadatan 2 × 105 dalam 100 μL media standar dengan 10 mg·mL −1 doksisiklin dan diinkubasi selama 24 jam. PBS ditambahkan pada sumur di sekitarnya untuk mencegah penguapan media. Media tersebut kemudian disedot, dan 100 μL obat yang diencerkan dengan 10 mg·mL −1 doksisiklin ditambahkan ke sumur yang sesuai dan diinkubasi selama 72 jam. Uji MTS (CellTiter 96 ® Promega) kemudian dilakukan seperti di atas untuk mengukur jumlah sel yang bertahan hidup. Hasil dianalisis menggunakan GraphPad Prism. Regresi nonlinier digunakan untuk menghitung nilai IC50.
2.8 Radiasi
Untuk menilai efek radiasi pada sel-sel yang mengekspresikan CHRDL2 secara berlebihan, sel-sel RKO ditanam pada kepadatan 1 × 105 dalam cawan bundar 10 cm dan diobati dengan kontrol DMSO atau pengobatan doksisiklin CHRDL2++. Setelah 24 jam, sel-sel diiradiasi menggunakan iradiasi sinar-X pada 0 GY, 2 GY, 4 GY dan 6 GY. Media dan pengobatan CHRDL2++ disegarkan, dan sel-sel diinkubasi dalam kondisi standar selama 48 jam berikutnya. Selanjutnya, jumlah sel dihitung menggunakan uji viabilitas sel Trypan biru (T10282, Thermo Scientific, Fisher Scientific).
2.9 Imunofluoresensi
Untuk deteksi protein seluler, sel-sel diletakkan pada penutup kaca dan ditumbuhkan hingga ~70% konfluensi. Sel-sel difiksasi dengan metanol, dan membran sel dipermeabilisasi dengan TRITON × 0,5% selama 5 menit. Sel-sel diblokir dengan 1% BSA selama 1 jam pada suhu 37 °C, lalu diinkubasi dengan antibodi primer dalam 1% BSA selama 1 jam pada suhu 37 °C. Antibodi sekunder kemudian ditambahkan ke sel-sel selama 1 jam pada suhu 37 °C. 5 μL media pemasangan dengan DAPI (VECTASHIELD® WZ-93952-27; Cole-Parmer, St. Neots, Inggris) kemudian ditempatkan pada penutup kaca, dan penutup kaca difiksasi pada slide untuk pencitraan. Gambar diperoleh menggunakan sistem Leica DM4000, dan fluoresensi sel total terkoreksi diperoleh menggunakan Imag J. Antibodi berikut digunakan: H2AX (1:50; Nomor katalog: ab195188, Abcam), ATM (1:50; Nomor katalog: ab2354, Abcam), P53 (1:800. Nomor katalog: 25275, Cell Signaling), B-catenin (1:100. Nomor katalog: 610154, BD biosciences, CA, AS), Ki67 (1:100. Nomor katalog: D3B5, Cell signaling), Rad21 (1:100. Nomor katalog: ab992, Abcam), Ku70 (1:100. Nomor katalog: 2172, Abcam), Alexa Fluor Goat anti-mouse AF488 (1:50. Nomor katalog: A-11001, Abcam) dan Alexa Fluor Kambing antikelinci AF568 (1:50. Nomor katalog: A-11011, Abcam).
2.10 Uji komet
Sel COLO320 ditanam pada kepadatan 1 × 105 dengan kontrol DMSO atau CHRDL2++ doksisiklin dan diobati dengan IC50 oksaliplatin. Setelah 72 jam, sel dipanen dan diencerkan dalam PBS pada kepadatan 104 / mL. Suspensi sel dicampur 1:5 dengan agarosa pembentuk gel rendah, dan 100 μL ditempatkan pada slide berlapis Polylysine yang dicelupkan dalam agarosa 1% (Merck) dan dibiarkan mengeras. 100 μL agarosa pembentuk gel rendah (Merck) lebih lanjut ditempatkan di atas dan dibiarkan mengeras. Lisis sel kemudian dilakukan dengan merendam slide dalam buffer lisis (garam natrium N-lauroylsarcosine 2% (Merck), 0,5 M NA 2 EDTA (Merck) dan 0,1 mg·mL −1 proteinase K) selama 1 jam. Kemudian slide dicuci dalam buffer elektroforesis selama 1,5 jam (90 mm Tris Buffer (Fisher Fisher Scientific, Loughborough, Inggris), 90 mm asam borat (Merck) dan 2 mm NA 2 EDTA ). Slide ditempatkan dalam tangki elektroforesis dan direndam dalam buffer elektroforesis, di bawah arus 20 V selama 40 menit. Slide kemudian diwarnai dengan 1% SYBR SAFE (Invitrogen, Fisher Scientific) dalam TBE selama 20 menit, sebelum dehidrasi melalui perendaman dalam etanol 70%, 90% dan 100%. Slide divisualisasikan oleh sistem Leica DM4000, dan ekor diukur menggunakan Image J.
2.11 Persiapan, kultur dan pemeliharaan organoid
Organoid dipelihara dalam atmosfer yang dilembabkan pada suhu 37 °C dengan 4% CO2. Organoid ditumbuhkan dalam media ADF seperti yang dijelaskan: Advanced DMEM/F12, 2 m m GLUTAMAX, 1 m m N-acetylcysteine dan 10 m m HEPES, yang dilengkapi dengan 1% PS, 10% B27 dan 5% N2. Faktor pertumbuhan juga diberikan ke media di sekitar ekstrak membran dasar (Cultrex, Bio-Techne, Minneapolis, MN, AS): 1% Noggin rekombinan tikus (PeproTech, London, Inggris), 1% EGF rekombinan tikus (Invitrogen, Fisher Scientific) dan 5% R-spondin manusia rekombinan (PeproTech, London, Inggris).
Organoid dihasilkan dari tikus tipe liar (C57BL6/J, diperoleh dari Charles River, Cambridge, Inggris) dari kedua jenis kelamin berusia antara 6 dan 12 minggu. Tikus ditempatkan bersama dalam kandang berventilasi individual yang dilengkapi dengan alas tidur, pengayaan, dan akses ad libitum ke makanan dan air. Semua prosedur dilakukan sesuai dengan ‘Animals (Scientific Procedures) Act 1986’ dari UK Home Office dan dengan persetujuan dari Brunel University Animals Welfare and Ethical Review Board dan di bawah lisensi pribadi, proyek, dan pendirian I0578EED1, PDF0B94C3, dan XEC0493FD. Secara singkat, kripta diisolasi dari usus halus tikus dan dicuci dengan PBS. Vili dan sel yang berdiferensiasi dikikis dari usus menggunakan slide mikroskop kaca. Bagian usus dipotong menjadi segmen 2 mm dan dipindahkan ke PBS dingin. Pipet dilapisi dengan FBS, dan segmen usus dicuci dengan pipet ke atas dan ke bawah untuk melepaskan sel tunggal dan serpihan. PBS dibuang, dan pencucian diulang 5 kali. Segmen kemudian disuspensikan kembali dalam EDTA/PBS 2,5 m m untuk melonggarkan kripta dan diputar pada suhu 4 °C selama 30 menit. Supernatan kemudian dibuang, dan segmen disuspensikan kembali dalam media ADF. Seluruh volume dipipet ke atas dan ke bawah beberapa kali, dan kemudian, supernatan dibuang dan disentrifugasi selama 5 menit pada 1200 rpm pada suhu 4 °C. Supernatan dibuang, dan pelet yang dihasilkan disuspensikan kembali dalam media ADF 10 mL dan dilewatkan melalui saringan sel 70 μm ke dalam tabung falcon 15 mL yang bersih. Tabung kemudian disentrifugasi selama 2 menit pada 600 rcf pada suhu 4 °C sehingga sel tunggal tidak akan masuk ke dalam pelet, dan supernatan dibuang. Ini diulangi sebanyak 3 kali. Akhirnya, pelet disuspensikan kembali dalam media ADF 50 μL dan Cultrex 100 μL, dan dipipet 40 μL/sumur. Pengaliran organoid diulang setiap 48 jam dan terdiri dari pemindahan organoid ke dalam tabung kerucut 15 mL, pemipetan ke atas dan ke bawah untuk memecah organoid. Organoid kemudian disentrifugasi selama 2 menit pada 600–800 rpm pada suhu 4 °C dan kemudian disuspensikan kembali dalam ADF dengan Cultrex seperti yang dijelaskan sebelumnya.
2.12 Perawatan kemoterapi organoid
Organoid ditanam dalam rangkap tiga di dalam pelat 24-sumur dan diberi perlakuan 5FU pada 0,5, 1, dan 5 μ m 5FU. Setelah 96 jam, gambar diambil dari setiap sumur, dan jumlah organoid hidup dihitung secara buta oleh peneliti independen.
2.13 Pewarnaan imunofluoresensi organoid
Sampel organoid disiapkan untuk pewarnaan dengan membuang media pertumbuhan dan dipeletkan melalui sentrifugasi pada 600 g . Organoid kemudian difiksasi melalui resuspensi dalam 500 μL formalin penyangga netral (Merck) selama 10 menit, sebelum dipeletkan pada 400 g dan resuspensi dalam etanol 70% selama 1 menit. Organoid kemudian dipeletkan pada 400 g dan disuspensi kembali dalam 50 μL agarosa pembentuk gel rendah (Merck) dan diinkubasi di atas es selama 30 menit, sebelum ditanamkan dalam blok parafin menggunakan protokol standar. Pemotongan organoid dilakukan pada 5 μm melalui pemotongan mikrotom standar dan dibiarkan mengering pada slide.
Kaca film yang berisi potongan organoid dihilangkan lilinnya melalui perendaman xylene (Fisher Scientific, Loughborough, Inggris) selama 5 menit dan dihidrasi ulang melalui perendaman dalam etanol pada 100%, 90% dan 70% selama 5 menit. Pengambilan antigen dilakukan dengan merendam kaca film dalam buffer natrium sitrat 10 mm yang mendidih ( Merck), sebelum dicuci dengan PBS. Sampel kemudian diblokir melalui penambahan serum Kambing (Zytochem Plus, 2bscientific, Kirtlington, Inggris) selama 1 jam. Antibodi primer yang diencerkan dalam PBS ditambahkan selama 1 jam, dan antibodi sekunder diinkubasi selama 1 jam dalam gelap. Penutup kaca film dipasang menggunakan VECTASHIELD Vibrance [TM] Antifade Mounting Medium dengan DAPI (2bscientific, Kirtlington, Inggris) untuk pencitraan. Kaca film divisualisasikan oleh sistem Leica DM4000. Pewarnaan organoid dinilai pada skala 1–5 oleh peneliti independen yang dibutakan. Antibodi berikut digunakan: OLFM4 (1:200; Nomor katalog: 39141, Cell Signalling), Alexa Flour Goat anti-kelinci AF568 (1:50; Nomor katalog: A-11011, Abcam) dan B-catenin (1:100; Nomor katalog: 610154, BD Biosciences, CA, AS).
2.14 RNA-sekuen
Sampel untuk analisis RNA-seq disiapkan dengan membudidayakan sel dalam kondisi media standar dengan ekspresi berlebih gen CHRDL2 melalui ekspresi yang dapat diinduksi doksisiklin. Doksisiklin diberikan dalam jumlah 10 μg·mL −1 , 1 μg ·mL −1 dan 0,1 μg ·mL −1 atau kontrol DMSO seperti yang dijelaskan sebelumnya.
RNA-seq dilakukan oleh Oxford Genomic Centre. Data untuk analisis bioinformatika diberikan dalam format pembacaan mentah fastq. Data dianalisis menggunakan paket perangkat lunak sumber terbuka Tuxedo Suite. Tophat2 dan Bowtie2 digunakan untuk memetakan pembacaan berpasangan ke genom Homo sapiens acuan GRCh38. GENCODE38 digunakan sebagai anotasi genom manusia acuan.
Hasil pembacaan yang selaras disaring berdasarkan kualitas menggunakan Samtools dengan ambang batas pemilihan minimum 30. Perakitan dan kuantifikasi transkrip dilakukan melalui Cufflinks, dan analisis ekspresi diferensial dicapai melalui penggunaan perangkat lunak Cuffdiff. Ekspresi diferensial dinyatakan dalam bentuk perubahan log 2 kali lipat antara sampel dan kontrol.
2.15 Visualisasi data dan R
Data dibersihkan, dan data signifikan diekstraksi menggunakan perangkat lunak R. Grafik dibuat menggunakan R studio 4.1.0 menggunakan pustaka ggplot2 dan heatmap2 .
Analisis pengayaan set gen dilakukan menggunakan perangkat lunak GSEA 4.2.3. Platform anotasi Chip yang digunakan adalah Human_Ensembl_Transcript_ID_MSigDB.v7.5.1.chip.
Set gen yang digunakan:
c6.all.v7.5.1.simbol.gmt
h.all.v7.5.1.simbol.gmt
PERATURAN GOBP TENTANG JALUR PENSINYALAN BMP
enplot_REAKSI_SINYAL_PI3K_AKT_PADA_KANKER_13
enplot_GOMF_BMP_PENERIMA_IKAT_58
WP_NRF2_PATHWAY.v2023.1.Hs.
3 Hasil
3.1 Ekspresi berlebihan CHRDL2 mengurangi proliferasi sel dan meningkatkan migrasi melalui aktivasi WNT
Untuk menentukan efek CHRDL2 pada sel kanker kolorektal, kami mentransduksi empat lini sel CRC yang dikarakterisasi secara ekstensif dengan sistem overekspresi yang diinduksi doksisiklin yang dikemas secara viral untuk cDNA CHRDL2 dengan panjang penuh . Lini sel adenokarsinoma kolorektal sengaja dipilih untuk mencakup berbagai tingkat ekspresi CHRDL2 dan BMP, serta mutasi genetik: CACO2 dan LS180 (CHRDL2 sedang), COLO320 dan RKO (sangat rendah) (Gbr. S1A–C ). Doksisiklin diberikan dalam 3 konsentrasi ke lini sel: 0,1 μg·mL −1 (CHRDL2), 1 μg·mL −1 (CHRDL2+) atau 10 μg·mL −1 (CHRDL2++) (Gbr. 1A ) untuk menginduksi ekspresi. qPCR dan western blotting mengonfirmasikan overekspresi CHRDL2 pada tingkat RNA dan protein, masing-masing (Gbr. 1B,C , Gbr. S1D ). Antagonisme BMP ditunjukkan melalui penilaian tingkat SMAD 1/5 terfosforilasi (Gbr. 1D , Gbr. S1E ). Pita dinormalisasi ke tingkat total SMAD1. Media terkondisi dari lini sel yang mengekspresikan CHRDL2 secara berlebihan juga dikumpulkan, dan protein CHRDL2 yang disekresikan ditemukan hadir di media (Gbr. 1B , Gbr. S1F ).
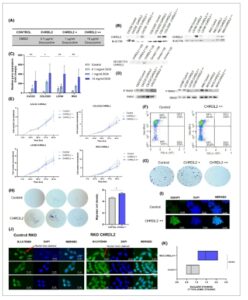
Gbr. 1
Buka di penampil gambar
Presentasi PowerPoint
Ekspresi berlebihan CHRDL2 yang dapat diinduksi pada lini sel kanker kolorektal (CRC) mengubah proliferasi dan migrasi. (A) Tabel pengobatan doksisiklin yang digunakan: sel kontrol diobati dengan dimetil sulfoksida (DMSO), sel uji diobati dengan doksisiklin pada: 0,1 μg·mL −1 (CHRDL2), 1 μg·mL −1 (CHRDL2+) atau 10 μg·mL −1 (CHRDL2++) untuk menginduksi ekspresi dari transgen CHRDL2. (B) Western blotting dari kadar protein CHRDL2 yang sesuai pada lini sel dengan ekspresi berlebihan lentivirus, dan CHRDL2 yang disekresikan yang terdapat dalam media kultur sel. Gambar representatif, N = 3. (C) qPCR kadar mRNA CHRDL2 yang diekspresikan sebagai perubahan lipat dalam 4 lini sel eksperimental. Garis sel ditumbuhkan dengan doksisiklin pada: 0,1, 1 atau 10 μg·mL −1 untuk menginduksi ekspresi. Uji- T ; RKO DMSO-10 μg·mL −1 P < 0,01, COLO320 DMSO-10 μg·mL −1 P < 0,05, CACO2 DMSO-10 μg·mL −1 P < 0,01, LS180 DMSO-10 μg·mL −1 P < 0,01. N = 3, uji- t . (D) Western blotting fosforilasi SMAD1/5 pada garis sel yang mengekspresikan CHRDL2 secara berlebihan. Gambar representatif, N = 3. (E) Uji MTT ((3-(4,5-dimetiltiazol-2-il)-2,5-difeniltetrazolium bromida)) terhadap proliferasi sel garis sel CHRDL2. Pada sel COLO320, ANOVA dua arah antara Kontrol dan CHRDL2++ adalah P < 0,0118. Sel LS180 ANOVA dua arah (analisis varians) antara Kontrol dan CHRDL2 = P < 0,0107. Kontrol dan CHRDL2++ P < 0,0114. Sel RKO Kontrol dan CHRDL2++ P < 0,0181. ANOVA satu arah pada 96 jam juga dilakukan: CACO2 P < 0,44, COLO320 P < 0,0411, LS180 P < 0,121, RKO P < 0,0476. N = 3. (F) Analisis flow cytometry sel COLO320 yang diberi overekspresi CHRDL2++ menggunakan pewarnaan antibodi Ki67 sebagai penanda sel yang berproliferasi. Proporsi sel Ki67 menurun dengan overekspresi CHRDL2++. Gambar representatif N = 3. (G) Pewarnaan kristal violet koloni sel RKO yang diobati dengan 1 μg·mL −1 dan 10 μg·mL −1 doksisiklin untuk menginduksi ekspresi CHRDL2. Gambar representatif N = 3. (H) Pewarnaan kristal violet sel COLO320 yang bermigrasi dengan 10 μg·mL −1 doksisiklin untuk menginduksi ekspresi CHRDL2. Kuantifikasi pada Gambar J menunjukkan peningkatan signifikan dalam jumlah sel yang bermigrasi dengan CHRD2++, t -test P < 0,0449. N = 3. (I) Pewarnaan imunofluoresensi IQGAP1 pada sel COLO320 dengan 10 μg·mL −1 doksisiklin untuk menginduksi ekspresi CHRDL2. N = 3. (J) Pewarnaan imunofluoresensi B-catenin pada sel RKO dengan 10 μg·mL −1 doksisiklin untuk menginduksi ekspresi CHRDL2. N = 3. (K) Kuantifikasi pewarnaan nuklir B-catenin pada pewarnaan sitoplasma pada sel RKO dengan 10 μg·mL −1 doksisiklin untuk menginduksi ekspresi CHRDL2 N = 3. Uji- T P < 0,0001. Di semua panel * P < 0,05, ** P < 0,01, **** P < 0,0001, ns, P > 0,05. Batang galat diberikan sebagai ± SEM. Skala batang menunjukkan 25 μ m .
Peningkatan proliferasi merupakan ciri khas sel kanker. Oleh karena itu, kami mengukur efek ekspresi berlebihan CHRDL2 pada proliferasi sel dalam lini sel kami. Seperti yang terlihat pada Gambar 1E , pertumbuhan sel sedikit berkurang selama ekspresi berlebihan CHRDL2 pada LS180 dan RKO ( P < 0,0114, P < 0,0181). Seperti yang terlihat pada Gambar 1F , ekspresi berlebihan CHRDL2 menurunkan proporsi sel yang berproliferasi, ditandai oleh Ki67+ (Gambar S2A , P < 0,05). Hal ini selanjutnya didukung oleh penurunan pewarnaan imunofluoresensi Ki67 ( P < 0,005) (Gambar S2B,C ). Pada populasi Ki67+, CHRDL2 meningkatkan jumlah sel fase S dan menurunkan proporsi dalam fase G2, mungkin mencerminkan penurunan laju proliferasi (Gambar S2D ). Investigasi kompetensi pembentukan koloni (klonogenisitas) sel-sel yang mengekspresikan CHRDL2 secara berlebihan menunjukkan bahwa potensi klonogenik berkurang (Gbr. 1G ). Hal ini ditemukan pada keempat lini sel yang diuji (Gbr. S2E , P < 0,01). Secara keseluruhan, bertentangan dengan hipotesis kami, ekspresi berlebihan CHRDL2 tampaknya mengurangi proliferasi dan pembentukan koloni.
Kemampuan migrasi adalah ukuran lebih lanjut dari kompetensi sel punca, jadi untuk mengukurnya dalam sel dengan ekspresi berlebihan CHRDL2, sel COLO320 disemai dalam sisipan transwell. Jumlah sel yang bermigrasi melalui membran berpori ke ruang bawah dihitung. Seperti yang terlihat pada Gambar 1H , ekspresi berlebihan CHRDL2 secara signifikan meningkatkan jumlah sel yang bermigrasi ( P <0,0449), yang menunjukkan peningkatan kemampuan migrasi, ciri khas sel punca kanker. Hal ini didukung oleh peningkatan ekspresi IQGAP1 P <0,0005 (Gambar 1I , Gambar S2F ), penanda perlekatan yang dikaitkan dengan kompetensi metastasis sel kanker [ [ 31 ] ].
Berikutnya, kami menyelidiki apakah ekspresi CHRDL2 meningkatkan kecenderungan sel epitel untuk mendapatkan kualitas seperti sel induk melalui peningkatan regulasi sinyal WNT melalui penghambatan BMP. Peningkatan sinyal WNT ditunjukkan dengan peningkatan pewarnaan B-catenin dalam nukleus dibandingkan pewarnaan sitoplasma. Seperti yang terlihat pada Gambar 1J,K , ekspresi berlebihan CHRDL2 meningkatkan lokalisasi nukleus B-catenin, ciri khas sinyal WNT ( P < 0,005, Gambar 1K ), yang mengonfirmasi hipotesis bahwa CHRDL2 meningkatkan sinyal WNT melalui penghambatan BMP.
Data ini, bersama dengan pengurangan fosforilasi SMAD, mengonfirmasi CHRDL2 sebagai antagonis BMP dan karenanya meningkatkan pensinyalan WNT seluler. Hal ini tampaknya menghasilkan peningkatan kemampuan migrasi, namun, juga mendukung fenotipe seluler yang tumbuh lebih lambat.
3.2 CHRDL2 meningkatkan resistensi terhadap kemoterapi umum
Karakteristik lain dari sel induk kanker adalah resistensinya terhadap kemoterapi [ [ 32 ] ]. Berdasarkan hal ini, penelitian kami bertujuan untuk mengevaluasi respons lini sel eksperimental kami terhadap tiga agen kemoterapi paling umum yang digunakan dalam pengobatan CRC.
Kami mengobati sel CHRDL2++ dengan obat kemoterapi, 5-Fluorouracil (5FU), irinotecan dan oxaliplatin dan menilai respons seluler melalui uji MTS. Gambar 2A menunjukkan pengurangan jumlah sel dengan peningkatan konsentrasi kemoterapi (μm ) . Lini sel kontrol diplot bersama dengan lini sel dengan ekspresi berlebihan CHRDL2, dan nilai setengah konsentrasi penghambatan maksimal (IC50) dihitung (Gbr. 2A,B ; Gbr. S3 ). Ekspresi berlebihan CHRDL2 secara signifikan meningkatkan resistensi terhadap kemoterapi di semua lini sel ( P < 0,01) seperti yang ditunjukkan oleh nilai IC50 yang meningkat (Gbr. S3 ). Peningkatan rata-rata nilai IC50 selama ekspresi berlebihan CHRDL2 untuk setiap obat dan lini sel dapat diamati pada Gbr. 2B . Peningkatan kelangsungan hidup terbesar (ditunjukkan oleh rasio IC50) terlihat pada sel COLO320 yang diobati dengan oxaliplatin, yang memiliki peningkatan 3,6 kali lipat. Sel yang diobati dengan nilai IC50 5FU mengalami penurunan ekspresi P-SMAD1/ (Gbr. 2C ), dibandingkan dengan sel yang tidak diobati dengan ekspresi berlebihan CHRDL2 (Gbr. 1D ). Hal ini menunjukkan bahwa sel dengan ekspresi CHRDL2 tertinggi mampu bertahan hidup dari kemoterapi (Gbr. 2C , Gbr. S4A ).
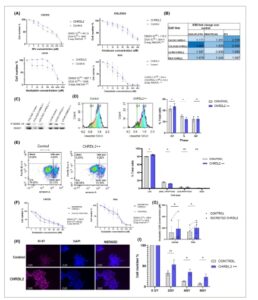
Gambar 2
Buka di penampil gambar
Presentasi PowerPoint
Peningkatan ekspresi CHRDL2 meningkatkan resistensi terhadap kemoterapi kanker kolorektal (CRC) umum. (A) Kurva dosis-respons obat menggunakan sel CACO2 dan 5-fluorouracil (5FU), sel COLO320 dan Irinotecan, serta sel LS180 dan RKO dan oxaliplatin N = 3. ANOVA dua arah (analisis varians) digunakan untuk menemukan perbedaan antar kurva, masing-masing P < 0,0068, P < 0,0001, P < 0,0006, P < 0,005. (B) Tabel rasio perbedaan nilai IC50 antara sel yang diobati dengan 10 μg·mL −1 doksisiklin (CHRDL2++) dan kontrol untuk setiap obat kemoterapi dan lini sel. N = 3. (C) Western blotting fosforilasi SMAD1/5 pada lini sel yang mengekspresikan CHRDL2 secara berlebihan melalui penambahan doksisiklin pada 1 μg·mL −1 (CHRDL2+) atau 10 μg·mL −1 (CHRDL2++) dan diobati dengan 10 μ m 5FU. Gambar representatif N = 3. (D) Analisis flow cytometry sel COLO320 yang diobati dengan oxaliplatin dan ekspresi berlebihan CHRDL2. CHRDL2 meningkatkan jumlah sel dalam fase G1. ( uji- t P < 0,003), dan mengurangi sel dalam fase S ( uji- t , P < 0,05) N = 3. (E) Analisis flow cytometry sel COLO320 yang diobati dengan oxaliplatin dan ekspresi berlebihan CHRDL2 diwarnai dengan Annexin-5 sebagai penanda apoptosis. Kuantifikasi persentase sel hidup, apoptotik, dan mati pada sel COLO320 yang diobati dengan oksaliplatin dan diberi ekspresi berlebih CHRDL2. CHRDL2 meningkatkan % sel hidup ( uji- t P < 0,03) dan menurunkan % sel apoptotik awal ( P < 0,023). N = 3. (F) Kurva dosis-respons obat sel CACO2 dan RKO dengan media terkondisi CHRDL2 dan oksaliplatin N = 3. ANOVA dua arah digunakan untuk menemukan perbedaan antar kurva, P < 0,005, P < 0,0001. (G) Nilai IC50 rata-rata untuk obat kemoterapi oksaliplatin, pada lini sel CACO2 dan RKO dengan media terkondisi CHRDL2. Media terkondisi dipanen dari lini sel yang sesuai dan diberikan ke lini sel parental untuk meniru pensinyalan parakrin CHRDL2 pada tingkat yang sama dari sistem ekspresi yang dapat diinduksi doksisiklin. CHRDL2 terhadap kontrol uji- t P < 0,0305. N = 3. (H) Pewarnaan imunofluoresensi Ki67 pada sel COLO320 yang diobati dengan 5 μ m oxaliplatin. (I) Jumlah sel setelah penyinaran sel RKO yang mengekspresikan CHRDL2++ secara berlebihan N = 3. Uji -t 4GY: P < 0,038, 6GY: P < 0,0241. Di semua panel * P < 0,05, ns, P > 0,05. Batang kesalahan diberikan sebagai ± SEM. Batang skala menunjukkan 25 μ m .
Sitometri aliran sel COLO320 yang diobati dengan oksaliplatin (Gbr. 2D ) menunjukkan peningkatan yang jelas dalam jumlah sel dalam fase S (baik pada sel kontrol maupun sel CHRDL2++) dibandingkan dengan sel yang tidak diobati (Gbr. S2D ). Hal ini mungkin disebabkan oleh terhentinya garpu replikasi akibat kerusakan DNA yang diberikan oleh kemoterapi dan aktivasi titik pemeriksaan S/G2. Menariknya, sel dengan ekspresi berlebihan CHRDL2 menunjukkan peningkatan yang lebih kecil pada sel yang terhenti di Fase S dibandingkan dengan kontrol dan oleh karena itu menunjukkan proporsi yang lebih besar yang hadir dalam fase G1. Ini adalah kebalikan dari sel yang tidak diobati, di mana CHRDL2 meningkatkan jumlah sel dalam Fase S yang dimungkinkan karena pembelahan sel yang lebih lambat. Analisis sitometri aliran lebih lanjut setelah pengobatan kemoterapi mengungkapkan bahwa ekspresi berlebihan CHRDL2 menurunkan jumlah sel yang telah memasuki apoptosis dini ( P < 0,05) (Gbr. 2E ) yang menunjukkan bahwa sel yang mengekspresikan berlebihan CHRDL2 memiliki kemampuan untuk menghindari apoptosis.
CHRDL2 yang disekresikan dari media terkondisi juga digunakan pada lini sel induk kami yang tidak mengekspresikan CHRDL2 untuk menilai pensinyalan parakrin. Media terkondisikan dipanen dari lini sel yang sesuai dengan transgen CHRDL2 yang dapat diinduksi dan sel kontrol induk. Induksi CHRDL2 untuk menghasilkan media terkondisikan dilakukan dengan menggunakan konsentrasi dan durasi pengobatan doksisiklin yang sama seperti sel-sel pada Gambar 2A . Sekali lagi, CHRDL2 yang disekresikan meningkatkan kelangsungan hidup sel selama kemoterapi dengan cara yang sama seperti sistem ekspresi berlebihan CHRDL2 intraseluler kami, P < 0,005 dengan nilai IC50 yang lebih tinggi (Gbr. 2F,G ).
Untuk menganalisis lebih lanjut kemampuan bertahan hidup sel-sel yang mengekspresikan CHRDL2 secara berlebihan selama kemoterapi, pewarnaan Ki67 dilakukan. Seperti yang terlihat pada Gambar 2H , ekspresi berlebihan CHRDL2 secara signifikan meningkatkan jumlah sel yang berproliferasi selama pengobatan dengan dosis rendah (IC25) obat kemoterapi oxaliplatin. Hal ini didukung oleh analisis flow cytometry ( P < 0,0055, Gambar S4B ) dan peningkatan regulasi pada Ki67 ( P < 0,0064, Gambar S4C ).
Resistensi terhadap radiasi, bersama dengan resistensi kemoterapi, juga dikaitkan dengan kanker yang lebih agresif yang menghindari pengobatan. ICS telah terbukti memiliki peningkatan resistensi terhadap radiasi; oleh karena itu, kami berusaha mengukur efek ekspresi berlebihan CHRDL2 pada kelangsungan hidup sel selama radiasi sinar-X. Sel diobati dengan radiasi sinar-X 0 GY, 2 GY, 4 GY atau 6 GY, dan viabilitas sel dinilai. Seperti yang terlihat pada Gambar 2I , ekspresi berlebihan CHRDL2 meningkatkan kelangsungan hidup sel pada radiasi 4 GY dan 6 GY ( P < 0,03, P < 0,02).
3.3 Ekspresi berlebihan CHRDL2 menurunkan kerusakan DNA selama pengobatan kemoterapi
Selanjutnya, kami menyelidiki mekanisme yang digunakan CHRDL2 untuk meningkatkan kelangsungan hidup sel selama pengobatan kemoterapi. Agen kemoterapi oxaliplatin diketahui menyebabkan ikatan silang DNA intra-strand, yang mengakibatkan pemutusan untai ganda (DSB), penangkapan siklus sel, dan apoptosis [ [ 33 ] ]. Oleh karena itu, kuantifikasi DSB dalam sel yang diobati dengan oxaliplatin dilakukan pada sel COLO320 CHRDL2++ menggunakan pewarnaan imunofluoresensi γH2AX dan Ku70. Kuantifikasi protein perbaikan DNA ATM dan RAD21 juga dilakukan untuk menilai apakah CHRDL2 melindungi sel dari kerusakan DNA melalui peningkatan jalur perbaikan DNA.
Gambar 3A menunjukkan DSB dalam sel yang diobati dengan dosis rendah oksaliplatin (sekitar IC25) setelah 24, 48, dan 72 jam pascaperawatan. Kuantifikasi menggunakan ImageJ menunjukkan bahwa sel yang mengekspresikan CHRDL2 secara berlebihan memiliki fokus γH2AX yang secara signifikan lebih sedikit dibandingkan dengan kontrol pada setiap titik waktu (Gbr. 3B P < 0,01, uji- t ). Perbedaan ini meningkat secara signifikan pada 72 jam dibandingkan dengan 24 jam. Hal ini menunjukkan bahwa CHRDL2 tidak selalu melindungi sel dari kerusakan DNA tetapi bertindak untuk mempercepat perbaikan kerusakan DNA jika dibandingkan dengan sel kontrol. Demikian pula, setelah 72 jam terdapat penurunan keberadaan Ku70, yang mengikat DSB untuk memfasilitasi nonhomologous end-joining (NHEJ), dalam sel CHRDL2 COLO320 ( P < 0,0057, Gbr. 3C dan Gbr. S4D ). Hal ini semakin menegaskan pengurangan kerusakan DNA melalui peningkatan CHRDL2.

Gambar 3
Buka di penampil gambar
Presentasi PowerPoint
Ekspresi berlebihan CHRDL2 menurunkan kerusakan DNA selama perawatan kemoterapi dan meningkatkan ekspresi jalur perbaikan DNA. (A) Gambaran representatif imunofluoresensi γH2AX pada sel COLO320 yang diobati dengan 5 μ m oksaliplatin pada 24, 48, dan 72 jam. Fokus ditunjukkan oleh panah merah N = 3. (B) Kuantifikasi fokus γH2AX pada sel COLO320 yang mengekspresikan berlebihan CHRDL2 yang diobati dengan 5 μ m oksaliplatin pada 24, 48, dan 72 jam. Sel diobati dengan reagen kontrol DMSO (dimetil sulfoksida), atau 10 μg·mL −1 Doxycycline untuk menginduksi ekspresi berlebihan CHRDL2 (CHRDL2++). Uji- T ; 24 jam P < 0,0001, 48 jam P < 0,01, 72 jam P < 0,0001. N = 3. (C) Pewarnaan imunofluoresensi Ku 70 pada sel COLO320 yang diobati dengan 5 μ m oksaliplatin. Gambar representatif N = 3. (D) Pewarnaan imunofluoresensi ATM dan RAD21 pada sel COLO320 yang diobati dengan 5 μ m oksaliplatin. Gambar representatif N = 3. (E) Kuantifikasi pewarnaan ATM dan RAD21 pada sel COLO320. Imunofluoresensi diberikan sebagai Fluoresensi Sel Total Terkoreksi (CTCF). Sel diobati dengan reagen kontrol DMSO, atau doksisiklin untuk menginduksi ekspresi berlebihan CHRDL2++. Uji- T ; P < 0,0001 dan P < 0,0023, masing-masing. N = 3. (F) Uji komet sel RKO yang diobati dengan IC50 Oxaliplatin. Sel kemudian diobati dengan overekspresi CHRDL2 ++ atau gambar representatif kontrol N = 3. (G) Kuantifikasi uji komet, uji- t P < 0,0001. N = 3. Di semua panel ** P < 0,01, *** P < 0,001, **** P < 0,0001, ns, P > 0,05. Batang kesalahan diberikan sebagai ± SEM. Kuantifikasi dilakukan menggunakan ImageJ. Batang skala menunjukkan 25 μ m di panel A, C, dan D. Batang skala menunjukkan 75 μ m di panel F.
Hal ini didukung oleh Gambar 3D dan E , di mana kami mengamati peningkatan ATM dan RAD21 secara signifikan ( P < 0,0001) ( P < 0,0023) pada sel yang mengekspresikan CHRDL2 secara berlebihan dibandingkan dengan kontrol, yang menunjukkan peningkatan jalur respons kerusakan DSB di mana ATM berfungsi sebagai transduser utama. Lebih jauh, kami telah menunjukkan peningkatan ekspresi ERCC1 dan PCNA, yang berfungsi dalam perbaikan ikatan silang DNA dan pemutusan untai tunggal (SSB), serta ARTEMIS, yang terlibat dalam perbaikan DSB melalui NHEJ (Gambar S5 ). Hal ini menunjukkan peningkatan jalur perbaikan DNA secara global oleh ekspresi berlebihan CHRDL2 setelah pengobatan kemoterapi.
Penanda DSB γH2AX juga diketahui terakumulasi selama penuaan sel. Akan tetapi, karena kami tidak menemukan perbedaan dalam ekspresi P53 (penanda penuaan, Gambar S5 ) pada sel-sel yang mengekspresikan CHRDL2 secara berlebihan, kemungkinan besar peningkatan jalur kerusakan DNA pada sel-sel CHRDL2++ melindungi terhadap kerusakan DNA akibat kemoterapi.
Kami selanjutnya menunjukkan kemampuan ekspresi berlebihan CHRDL2 untuk mengurangi kerusakan DNA selama kemoterapi dengan uji komet alkali, seperti yang diamati pada Gambar 3F dan G. Sel-sel diperlakukan dengan cara yang sama dengan oksaliplatin IC25. Kami mengamati sel-sel dengan ekspresi berlebihan CHRDL2 memiliki ‘ekor’ yang lebih pendek dari kometnya, yang menunjukkan DNA yang kurang terfragmentasi atau rusak. Kuantifikasi menggunakan ImageJ mengonfirmasi hal ini, dengan sel-sel CHRDL2++ memiliki panjang ekor yang berkurang secara signifikan dibandingkan dengan sel-sel kontrol ( P < 0,0001).
Di sini, kami telah menunjukkan bahwa ekspresi berlebihan CHRDL2 tidak hanya mengurangi kerusakan DNA selama kemoterapi, tetapi juga menghasilkan jalur perbaikan DNA yang aktif, sehingga membersihkan kerusakan DNA. Hal ini memungkinkan model sel kanker kami untuk meningkatkan kemampuan bertahan hidup selama kemoterapi dan radioterapi.
3.4 CHRDL2 menurunkan tunas organoid dan meningkatkan penanda sel induk
Lini sel CRC menunjukkan banyak kelainan, ditandai dengan banyaknya mutasi, peningkatan sinyal WNT, dan gangguan mekanisme perbaikan DNA. Oleh karena itu, kami juga meneliti dampak ekspresi berlebihan CHRDL2 pada ISC normal dalam model organoid, yang bertujuan untuk menjelaskan peran CHRDL2 dalam inisiasi tumor.
Untuk mereplikasi ceruk sel induk dan ekspresi berlebihan sinyal parakrin CHRDL2, yang biasanya berasal dari sel mesenkimal, kami membentuk organoid usus tikus, yang menyediakan platform tiga dimensi untuk memodelkan efek CHRDL2. Organoid ini terpapar pada bentuk CHRDL2 yang disekresikan dalam bentuk media terkondisi yang diperoleh dari lini sel kami. Sebagai kontrol, media terkondisi dari sel induk (yang tidak mengekspresikan CHRDL2) yang menjalani pengobatan doksisiklin juga dikumpulkan.
Organoid usus halus menunjukkan ciri morfologi yang berbeda, ditandai dengan konfigurasi seperti kuncup vili, dengan lapisan epitel luar membentuk tonjolan dan invaginasi yang berbeda. Penelitian sebelumnya telah meneliti profil ekspresi gen organoid usus, yang mengungkapkan bahwa sel epitel dalam ‘kuncup’ organoid menunjukkan pola ekspresi seperti kripta, sedangkan evaginasi menunjukkan ekspresi seperti vili [ [ 34 ] ].
Seperti yang terlihat pada Gambar 4A , setelah penambahan CHRDL2 ekstrinsik ke organoid, pengurangan yang nyata dalam jumlah kuncup yang berdiferensiasi diamati ( P < 0,001) (Gambar 4A,B ). Organoid mengembangkan karakteristik yang lebih kecil dan lebih melingkar, menunjukkan pertumbuhan yang lebih lambat mirip dengan pengamatan kami pada garis sel ( P < 0,001) (Gambar 4A,C ). Hal ini didukung oleh keberadaan olfactomedin-4 (OLFM4), penanda untuk sel induk LGR5+ dan target hilir pensinyalan WNT [ [ 35 ] ], yang meningkat pada organoid yang diobati dengan CHRDL2 ( P < 0,0010) (Gambar 4D,E ). Seperti yang terlihat pada Gambar 4F , dalam organoid kontrol, β-catenin terlokalisasi ke membran sel luar (panah biru), sedangkan pada perlakuan CHRDL2 ekstrinsik, β-catenin dapat diamati dalam sitoplasma dan nukleus sel organoid (panah merah). Selain itu, seperti yang diilustrasikan dalam Gambar 4G , kami mengamati peningkatan yang signifikan dibandingkan dengan kontrol ( P <0,05) dalam ekspresi penanda sel induk LGR5 (menunjukkan CBC kripta) dan BMI1 (sel induk kripta siklus lambat), dan juga SOX9 dan MSI1. Lebih jauh lagi, kami telah menunjukkan peningkatan pensinyalan WNT melalui peningkatan yang diamati dalam lokalisasi nuklir β-catenin.

Gambar 4
Buka di penampil gambar
Presentasi PowerPoint
CHRDL2 yang disekresikan menurunkan diferensiasi organoid usus halus tikus dan meningkatkan ekspresi penanda sel induk. (A) Gambar organoid turunan tikus yang diolah dengan media terkondisi yang mengandung bentuk CHRDL2 yang disekresikan dibandingkan dengan media terkondisi dari sel kontrol tanpa gambaran representatif ekspresi berlebihan CHRDL2 N = 3. (B) Kuantifikasi kuncup per organoid dalam organoid tikus yang diolah dengan CHRDL2 dibandingkan dengan kontrol. Uji- T P < 0,0001 N = 3. (C) Kuantifikasi diameter organoid rata-rata dalam organoid tikus yang diolah dengan CHRDL2 dibandingkan dengan kontrol. Uji- T P < 0,001 N = 3. (D) Pewarnaan imunofluoresensi protein OLFM4 pada organoid tikus yang diobati dengan CHRDL2 yang disekresikan dibandingkan dengan kontrol setelah gambar representatif 1 minggu N = 3. (E) Kuantifikasi penilaian imunofluoresensi OLFM4 pada organoid tikus yang diobati dengan CHRDL2 yang disekresikan dibandingkan dengan kontrol. Uji- T P < 0,0001 N = 3. (F) Pewarnaan imunofluoresensi lokalisasi β-catenin pada organoid usus halus tikus setelah penambahan CHRDL2 dan kontrol Gambar representatif N = 3. (G) qPCR penanda sel punca dari organoid tikus yang diobati dengan CHRDL2 dibandingkan dengan kontrol. T -test SOX9 P < 0,0014, LGR5 P < 0,04, P < 0,043, BMI1, P < 0,49, P < 0,0113, MSI1 P < 0,0067, P < 0,009. N = 3. (H) Gambar organoid turunan murine yang diobati dengan media terkondisikan yang mengandung bentuk CHRDL2 yang disekresikan dibandingkan dengan kontrol dan 5-fluorouracil (5FU). Gambar yang diambil 96 jam pasca-perawatan merupakan gambar representatif N = 3. (I) Kuantifikasi jumlah organoid hidup dalam organoid yang diobati dengan CHRDL2 dibandingkan dengan kontrol. N = 3. ANOVA dua arah yang membandingkan jumlah organoid hidup dalam kelompok kontrol dan kelompok yang diobati dengan CHRDL2 pada semua konsentrasi obat, P < 0,0442. Pada semua panel * P < 0,05, ** P < 0,01, *** P < 0,001, **** P < 0,0001, ns, P > 0,05. Batang kesalahan diberikan sebagai ± SEM. Batang skala menunjukkan 1000, 200, 50 dan 40 μ m pada panel A. Batang skala menunjukkan 50 μ m pada panel D dan F. Batang skala menunjukkan 1000 μ m pada panel H.
Organoid yang diobati dengan CHRDL2 menunjukkan resistensi terhadap pengobatan kemoterapi, seperti yang diamati pada Gambar 4H,I . Pada 5 μ m 5FU, jumlah organoid kontrol berkurang hingga kurang dari setengah sampel yang tidak diobati sedangkan organoid yang diobati dengan CHRDL2 menunjukkan sedikit atau tidak ada kematian. Jika digabungkan dengan peningkatan penanda sel punca, temuan ini secara kolektif menunjukkan bahwa paparan organoid usus terhadap CHRDL2 mengurangi diferensiasi melalui peningkatan pensinyalan WNT dan meningkatkan jumlah sel punca, yang mengarah pada peningkatan resistensi kemoterapi.
3.5 CHRDL2 meningkatkan jalur sel induk kanker
Untuk menjelaskan jalur-jalur di mana overekspresi CHRDL2 bekerja, analisis RNA-seq pada sel-sel CACO2 yang diberi pengobatan CHRDL2+ dan CHRDL2++ dilakukan dengan sel-sel yang diobati dengan DMSO sebagai garis dasar. Analisis ekspresi diferensial dilakukan pada data RNA-seq (Gbr. 5A ). 76 dan 145 gen yang diekspresikan secara diferensial diidentifikasi dalam kelompok CHRDL2 + dan CHRDL2++, masing-masing, pada P < 0,05 (Gbr. 5B ). Dari sini, kami memilih 22 gen yang diekspresikan secara diferensial dalam CHRDL2+ dan CHRDL2++ vs sel kontrol untuk analisis hilir (Gbr. 5C ). qPCR digunakan untuk mengonfirmasi perubahan ekspresi dalam replikasi biologis di sel CACO2 dan COLO320 (Gbr. 5D ).

Gambar 5
Buka di penampil gambar
Presentasi PowerPoint
Analisis RNA-seq menunjukkan bahwa ekspresi CHRDL2 meningkatkan sel induk kanker dan jalur khas kanker lainnya. Analisis RNA-seq sel CACO2 yang diobati dengan 1 μg·mL −1 (CHRDL2+) atau 10 μg·mL −1 (CHRDL2++) dibandingkan dengan sel kontrol yang diobati dengan DMSO (dimetil sulfoksida) N = 3. (A) Plot gunung berapi gen yang diekspresikan secara berbeda dari sel dengan ekspresi berlebihan CHRDL2 (CHRDL2+ dan CHRDL2++) dari analisis RNA-seq. Dinyatakan sebagai perubahan Log 2 Fold terhadap kontrol. (B) Plot batang gen yang diekspresikan secara berbeda secara signifikan dalam sel CHRDL2+ dan CHRDL2++. Gen yang disertakan melewati ambang batas P < 0,001 untuk CHRDL2+ dan P < 0,001 untuk CHRDL2++. (C) Perpotongan gen yang diekspresikan secara sangat berbeda pada kedua kelompok yang diberi CHRDL2+ dan CHRDL2++ P < 0,05. 21 gen diekspresikan secara berbeda pada kedua kelompok. (D) Validasi qPCR dari gen yang diekspresikan secara berbeda dari data RNA-seq dalam sel CACO2 dan COLO320 N = 3. (E) Plot GSEA dari jalur yang diekspresikan secara berbeda dalam sel CACO2 yang diberi CHRDL2++. (F) Validasi qPCR dari penanda sel punca dalam sel CACO2 dan COLO320 N = 3. Uji- T ; CACO2 BMI1 P < 0,05, CACO2 LGR6 P < 0,05, COLO320 LGR5 P < 0,01. N = 3. Di semua panel * P < 0,05, ** P < 0,01, ns, P > 0,05. Batang galat diberikan sebagai ± SEM.
Analisis pengayaan set gen (GSEA) pada seluruh set data RNA-seq mengungkapkan peningkatan regulasi jalur pensinyalan WNT yang khas ( P < 0,001) dan pengayaan gen yang memodulasi frekuensi, laju atau luas aktivitas jalur pensinyalan reseptor BMP ( P < 0,05). Lebih jauh lagi, gen target spesifik BMP ID1, yang diinduksi setelah stimulasi BMP, diturunkan regulasinya dalam set data kami oleh ekspresi berlebihan CHRDL2. APOA4, APOB dan APOC3, yang telah terbukti diinduksi secara spesifik oleh BMP 2 dan 4, juga dikurangi oleh ekspresi berlebihan CHRDL2 [ [ 36 ] ]. Hal ini menunjukkan bahwa ekspresi berlebihan CHRDL2 menghasilkan pengurangan langsung pensinyalan BMP yang dimediasi oleh BMP 2 dan 4. Peningkatan pensinyalan WNT dan modulasi pensinyalan BMP ini (Gbr. 5E ) memverifikasi peran CHRDL2 sebagai antagonis BMP dalam sel kanker usus besar. Jalur pensinyalan MYC dan pensinyalan LEF1, yang merupakan transduser hilir pensinyalan WNT, juga meningkat.
Kami selanjutnya menyelidiki penanda sel punca LGR5 dan LGR6 , serta homolog daerah penyisipan 1 Mo-MLV limfoma B ( BMI1 ) dan menunjukkan peningkatan regulasi oleh QPCR, dengan LGR6 , transduser WNT, juga meningkat regulasinya dalam data RNA-seq dalam pengobatan CHRDL2++ (Gbr. 5F ). Kami mencatat bahwa jalur BMI1 juga disorot oleh GSEA (Gbr. S6 ), jalur penentu sel punca dan berkorelasi dengan data Q-PCR kami.
GSEA mengungkapkan peningkatan jalur ciri khas kanker, transisi epitel ke mesenkimal (EMT) ( P < 0,001), dan angiogenesis, yang sering kali meningkat pada kanker kolorektal metastatik (Gbr. S6 ). Jalur perbaikan DNA juga meningkat secara signifikan termasuk gen perbaikan DSB utama BRCA1, RAD51 dan RAD52, yang mendukung temuan kami sehubungan dengan resistensi kemoterapi/radioterapi ( P < 0,05). Ada juga peningkatan regulasi yang signifikan dari pensinyalan RAF dan MTOR, yang sering kali dimodulasi selama perkembangan kanker. Lebih jauh lagi, gen terkait siklus sel yang meningkat regulasinya oleh knockout Rb juga meningkat regulasinya oleh CHRDL2, yang menunjukkan peningkatan pensinyalan protein siklus sel (Gbr. S6 ). Analisis ontologi gen mendukung temuan ini dengan pengayaan dalam proses biologis seperti adhesi seluler, apoptosis dan diferensiasi (Gbr. S7 ).
Menariknya, beberapa gen yang terlibat dalam metabolisme dan stres oksidatif dideregulasi oleh CHRDL2, termasuk TXNIP dan ENOX1, yang memengaruhi spesies oksigen reaktif, ADM2 dan PCK2, yang terlibat dalam stres metabolik, dan PCK2 yang penting untuk metabolisme yang tidak bergantung pada glukosa. Analisis GSEA juga menunjukkan deregulasi dalam pensinyalan glikolisis dan pengurangan fosforilasi oksidatif.
Gen yang terlibat dalam invasi sel kanker, CTSK dan LOXL4, juga mengalami peningkatan regulasi, meningkatkan pengamatan kami pada penanda jalur EMT. Pengamatan fenotipik migrasi dan invasi lebih lanjut dapat disebabkan oleh stimulasi RAS/RAF/MEK/ERK karena peningkatan regulasi ANKFN1 dan EGR1 [ [ 37 ] ]. Sekali lagi, hal ini dikuatkan oleh analisis GSEA, yang menunjukkan pengayaan jalur pensinyalan RAF (Gbr. S7 ). Kami juga menemukan peningkatan regulasi MAPK10, yang telah terbukti menghambat apoptosis, yang terkait dengan peningkatan kemampuan bertahan hidup yang kami amati. Namun, kami juga melihat penurunan regulasi DUSP6, yang mengaktifkan MAPK10 yang terdefosforilasi.
Ada juga bukti deregulasi beberapa penanda jalur imun dan inflamasi oleh ekspresi berlebihan CHRDL2. IL15 terlibat dalam infiltrasi sel imun ke lokasi tumor, dan CEBP menstimulasi penanda inflamasi IL6. Pensinyalan IL2 juga meningkat dalam data GSEA kami, yang menunjukkan peningkatan infiltrasi imun (Gbr. S6 ). ITH1, yang telah terbukti menstabilkan ECM selama inflamasi, mengalami penurunan regulasi dalam data kami dan juga telah terbukti hilang dalam banyak jenis kanker. Ini menunjukkan peningkatan jalur imun dan inflamasi oleh ekspresi berlebihan CHRDL2.
Telah diamati adanya penurunan regulasi gen yang banyak diekspresikan dalam lapisan dan mukosa usus yang sehat, seperti NCALD dan TIFF1. NCALD, sensor kalsium, ditemukan banyak diekspresikan dalam saluran usus, begitu pula TIFF1, yang membantu pembentukan mukosa usus. Hal ini dapat menandakan dediferensiasi sel epitel kanker kita.
Singkatnya, gen dan jalur yang diekspresikan secara berbeda akibat peningkatan regulasi CHRDL2 mencakup pemeliharaan sel induk, perubahan metabolik, invasi, migrasi dan proses EMT, perbaikan DNA dan infiltrasi imun. Semua ini merupakan proses yang dikarakterisasi dengan baik dalam perkembangan kanker serta respons pengobatan, yang menyoroti CHRDL2 sebagai kandidat penting untuk studi biomarker dan target terapeutik lebih lanjut.
4 Diskusi
Peran pensinyalan BMP dalam kanker telah dipelajari dengan baik tetapi sering kali paradoks; dengan pensinyalan BMP terbukti diperlukan untuk mencegah pensinyalan WNT terkait kanker dan ISC keluar dari kripta intestinal [ [ 12 , 25 ] ]. Sebaliknya, ia juga dapat berperan dalam mendorong tumorigenesis [ [ 16 ] ]. Namun, jelas bahwa antagonis BMP memainkan peran fungsional penting dalam mengatur pensinyalan BMP dan oleh karena itu dapat menjadi biomarker utama dalam perkembangan kanker [ [ 20 ] ]. Dalam penelitian ini, kami telah mengonfirmasi bahwa CHRDL2 menekan pensinyalan BMP dalam sel CRC, yang menyebabkan peningkatan pensinyalan WNT, dan menyebabkan perubahan dalam pertumbuhan sel, respons terhadap kemoterapi, dan karakteristik sel punca.
Sebelumnya, CHRDL2 telah terbukti mengikat BMP2 dan 9 untuk memblokir sinyal fosforilasi SMAD1/5 yang dimediasi BMP [ [ 27 , 30 ] ]. Studi kelangsungan hidup pasien telah menunjukkan bahwa ekspresi berlebihan CHRDL2 memprediksi prognosis yang buruk pada pasien CRC, dan ekspresi mRNA meningkat pada jaringan tumor pasien dibandingkan dengan kontrol. Variasi di dekat CHRDL2 juga telah dikaitkan sebagai penyebab peningkatan risiko CRC dalam studi asosiasi genom-lebar (GWAS) [ [ 38 , 39 ] ]. Studi knockdown CHRDL2 telah terbukti menghambat proliferasi dan migrasi pada CRC, kanker lambung dan osteosarkoma, dan ekspresi berlebihan meningkatkan proliferasi seluler, migrasi dan klonogenisitas [ [ 27 , 30 ] ]. Karena antagonis BMP lain yang disekresikan, Noggin dan GREM1, juga terbukti meningkatkan tumorigenesis dan memodulasi sel induk usus, kami menyarankan bahwa CHRDL2 bekerja dengan cara yang sama dan ini melalui jalur BMP [ [ 21 , 23 , 25 ] ].
Di sini kami telah menggunakan model yang dapat diinduksi doksisiklin untuk mengekspresikan CHRDL2 secara berlebihan pada berbagai tingkat untuk menyelidiki efek transkripsi dan perilaku gen ini. Kami telah menunjukkan bahwa sebagai antagonis BMP, CHRDL2 mengurangi tingkat SMAD1/5/9 yang terfosforilasi, yang menunjukkan berkurangnya pensinyalan BMP. Selain itu, kami telah menunjukkan peningkatan pensinyalan WNT setelah penambahan CHRDL2 pada lini sel dan model organoid, yang ditunjukkan melalui lokalisasi β-catenin ke dalam nukleus.
Meskipun ada laporan sebelumnya tentang CHRDL2 yang meningkatkan proliferasi, kami menemukan sedikit penurunan proliferasi di lini sel CRC kami, yang tercermin dari berkurangnya sel Ki-67+ dan berkurangnya pertumbuhan koloni.
Kami juga mengamati peningkatan migrasi sel melalui membran berpori dari sel-sel yang mengekspresikan CHRDL2 secara berlebihan. CSC telah lama terbukti memiliki potensi migrasi yang meningkat, yang mendukung temuan kami tentang peningkatan kualitas seperti sel induk selama ekspresi berlebihan CHRDL2. Hal ini tercermin dalam data RNA-seq kami, yang menunjukkan pengayaan untuk jalur EMT, yang bergantung pada peningkatan sifat migrasi dan invasi sel kanker.
Berikutnya, kami mengidentifikasi peningkatan resistensi terhadap tiga bentuk kemoterapi yang paling umum digunakan untuk mengobati CRC, sekali lagi memperkuat kecenderungan untuk bertahan hidup selama ekspresi berlebihan CHRDL2 dalam model sel CRC kami. Standar perawatan untuk semua kecuali tahap awal CRC bergantung pada penggunaan kemoterapi dalam kombinasi dengan prosedur pembedahan. 5-Fluorouracil (5FU) saat ini merupakan landasan pengobatan kemoterapi yang digunakan untuk mengobati CRC [ [ 40 , 41 ] ] dan digunakan dalam kombinasi dengan Oxaliplatin, senyawa diamino cyclohexane platinum yang membentuk DNA addukt (dikenal sebagai FOLFOX) atau Irinotecan, penghambat topoisomerase I (dikenal sebagai FOLFIRI) [ [ 42 ] ]. Di setiap lini sel yang kami uji, ada peningkatan kelangsungan hidup selama pengobatan kemoterapi, terlepas dari mekanisme kerja dari tiga agen kemoterapi. Ini juga terlihat pada organoid yang diobati dengan CHRDL2 yang disekresikan, yang memiliki peningkatan kemampuan bertahan hidup setelah paparan 5FU dibandingkan dengan kontrol. Resistensi kemoterapi dikonfirmasi melalui analisis flow cytometry yang menunjukkan bahwa ekspresi berlebihan CHRDL2 mengurangi jumlah sel yang memasuki apoptosis dan juga meningkatkan jumlah sel yang berproliferasi yang tersisa setelah pengobatan. Selain itu, kami telah menunjukkan bahwa ekspresi berlebihan CHRDL2 meningkatkan kelangsungan hidup sel selama pengobatan iradiasi, yang dapat digunakan bersamaan dengan kemoterapi dalam pengobatan CRC.
Kami mengamati peningkatan regulasi beberapa protein perbaikan DNA utama, seperti sinyal RAD21, ATM, PCNA, ERCC1, ARTMEIS dan BRCA1 selama kemoterapi sel yang mengekspresikan CHRDL2 secara berlebihan yang menunjukkan jalur respons kerusakan DNA yang hiperaktif. Hal ini kemungkinan menjadi faktor dalam percepatan pembersihan DSB yang ditandai dengan pengurangan fokus γH2AX dan Ku70 yang jauh lebih cepat. Gen perbaikan kerusakan DNA juga terbukti mengalami peningkatan regulasi oleh data RNA-seq kami, seperti yang ditunjukkan oleh GSEA, meskipun sel-sel ini tidak terpapar kemoterapi apa pun, dan oleh karena itu, kami hanya mengharapkan aktivitas perbaikan DNA dasar. Jalur perbaikan mana yang mengalami peningkatan regulasi sebagai respons terhadap sel CHRDL2 yang menjalani kemoterapi, dan apakah ini rawan kesalahan atau akurat, merupakan pertanyaan penting yang masih harus dijawab. Secara umum, peningkatan aktivasi respons kerusakan DNA juga merupakan ciri khas CSC, yang telah terbukti secara luas membantu kelangsungan hidup CSC setelah pengobatan konvensional, memungkinkan kembalinya kanker pada pasien dan memperburuk prognosis [ [ 43 – 45 ] ].
Melalui analisis RNA-seq yang komprehensif dan validasi qPCR, kami telah menunjukkan bahwa ekspresi berlebihan CHRDL2 meningkatkan pensinyalan WNT dan ekspresi penanda sel punca, termasuk LGR5, BMI1, LGR6, dan SOX9 dalam model 2D dan 3D. Data ini secara kolektif menunjukkan bahwa CHRDL2 meningkatkan kapasitas sel punca melalui peningkatan pensinyalan WNT dan karenanya mengintensifkan fenotipe seperti sel punca.
Namun, CHRDL2 tidak meningkatkan proliferasi dan klonogenisitas, yang menunjukkan bahwa CHRDL2 lebih mendukung sel punca kanker yang siklusnya lebih lambat dibandingkan dengan sel punca kanker yang hiperproliferatif. Di dalam kripta intestinal, sel punca normal tersusun dalam hierarki, dengan sel punca yang berkembang biak dengan cepat atau sel kolumnar dasar kripta (CBC) di dasar kripta. Populasi sel punca yang terpisah terletak pada posisi +4, yang tampaknya siklusnya lebih lambat [ [ 4 , 5 ] ]. Meskipun datanya tidak konsisten, beberapa penelitian mengusulkan bahwa sel punca siklus lambat pada posisi +4 ini ditandai oleh BMI1, yang diatur naik dalam data RNA-seq dan qPCR kami, yang meningkatkan kemungkinan bahwa CHRDL2 meningkatkan fenotipe sel punca siklus lambat ini. CSC siklus lambat juga telah terbukti tahan terhadap radiasi, mirip dengan sel yang mengekspresikan CHRDL2 secara berlebihan [ [ 46 , 47 ] ]. Ada beberapa bukti yang saling bertentangan mengenai peran sel punca siklus lambat ini dengan sejumlah publikasi yang menyatakan bahwa sel-sel ini merupakan kunci regenerasi usus setelah cedera [ [ 5 , 48 ] ]. Namun, penelitian yang lebih baru menunjukkan bahwa LGR5+ CBC juga mampu memenuhi peran ini, atau menunjukkan bahwa kedua populasi ini saling mendukung untuk memfasilitasi perbaikan jaringan [ [ 6 ] ].
Model organoid kami yang diobati dengan CHRDL2 yang disekresikan selanjutnya mengonfirmasi hipotesis mirip sel punca kami yang meningkat. Ketika organoid tikus diobati dengan CHRDL2, mereka menunjukkan pengurangan yang signifikan dalam pembentukan tunas terdiferensiasi, menciptakan fenotipe mirip sel punca yang lebih kecil dan bulat dibandingkan dengan kontrol. Pewarnaan imunofluoresensi penanda sel punca OLFM4 menunjukkan peningkatan regulasi pada organoid yang diobati dengan CHRDL2. Lebih jauh, penanda sel punca LGR5 , MSI1 , BMI1 dan SOX9 meningkat setelah pengobatan CHRDL2, menunjukkan bahwa penghambatan pensinyalan BMP ekstraseluler dapat secara langsung meningkatkan keseluruh sel punca dalam sel usus normal. Dalam penelitian terbaru, antagonis BMP lain, GREM1, meningkat regulasinya dalam stroma usus sebagai respons terhadap cedera, menghasilkan pemrograman ulang/dedifferensiasi dalam sel epitel usus untuk mendorong perbaikan [ [ 49 ] ]. Sebaliknya, pengobatan organoid usus manusia dengan BMP, meskipun tidak menyebabkan perbedaan morfologi yang besar, mengakibatkan berkurangnya ekspresi penanda sel induk, termasuk OLFM4 [ [ 36 ] ]. Dengan demikian, penghambatan pensinyalan BMP melalui CHRDL2 atau antagonis lain mungkin memang memaksa sel kanker di usus besar ke dalam keadaan yang lebih mirip sel induk, dan meskipun ini mungkin tidak meningkatkan laju proliferasi, hal ini dapat meningkatkan umur panjang dan kelangsungan hidup sel-sel ini selama pengobatan dengan terapi yang merusak DNA.
Melalui analisis RNA-seq, kami juga telah menunjukkan ekspresi diferensial dari biomarker kanker lain oleh CHRDL2, seperti EGR1 , REG4 , dan TFF1 , yang telah terbukti mengatur proliferasi, migrasi, dan metastasis. Lebih jauh lagi, CHRDL2 ditemukan berdampak berbeda pada beberapa jalur kanker utama, termasuk jalur EMT, MYC, MTOR, PI3/AKT, dan RAF. Misalnya, DDIT4 adalah pengatur MTOR dan diatur naik oleh CHRDL2 dan EGR1, yang bekerja melalui PI3K/AKT, diatur turun oleh CHRDL2. Memang, CHRDL2 sebelumnya telah terbukti bekerja melalui PI3K/AKT pada osteosarkoma [ [ 30 ] ]. Sementara perubahan dalam pensinyalan BMP dan WNT yang ditunjukkan dalam analisis GSEA kami menunjukkan bahwa efek CHRDL2 dalam sistem kami bekerja secara langsung melalui penghambatan BMP, tidak mungkin untuk mengesampingkan bahwa beberapa jalur dipengaruhi oleh tindakan CHRLD2 yang tidak bergantung pada BMP. Memang, Wang dkk. menunjukkan bahwa CHRDL2 dapat secara langsung mengubah fosforilasi dan aktivitas YAP dalam lini sel kanker lambung, yang memerlukan eksplorasi lebih lanjut (Wang dkk., 2022). Data ini memberikan jalan baru untuk penelitian tentang mekanisme yang dapat digunakan CHRDL2 dan antagonis BMP lainnya untuk memberikan efeknya. Mengungkap jalur yang dimodulasi oleh CHRDL2 dan antagonis BMP lainnya tidak diragukan lagi akan mendorong penyelidikan masa depan dalam penelitian kanker.
5 Kesimpulan
Temuan kami menunjukkan bahwa CHRDL2 harus dieksplorasi lebih lanjut sebagai biomarker potensial untuk peningkatan resistensi kemoterapi pada CRC. Hal ini akan memerlukan pemahaman yang lebih mendalam tentang mekanisme resistensi dan juga penentuan pola ekspresi berlebihan CHRDL2 secara akurat selama tumorigenesis dan terapi. Singkatnya, data kami sangat menyarankan bahwa CHRDL2, dengan menghambat pensinyalan BMP dan menambah pensinyalan WNT, meningkatkan sifat sel punca dalam sel kanker, sehingga berkontribusi pada perkembangan kanker dan kemungkinan resistensi terapeutik.